Synthesis of high-quality libraries of long (150mer) oligonucleotides by a novel depurination controlled process
1
2010
... DNA的人工合成是合成生物学研究的底层推动技术.DNA化学合成法是当前主流的商业化合成方法,在合成长度和合成通量上不断取得突破,发展出柱式合成和芯片合成工艺,使寡核苷酸链合成通量可以达到百万条级,合成成本降低约3个数量级[1].近年来,DNA酶促合成法日益受到关注,利用末端脱氧核苷酸转移酶(TdT)在温和的条件进行寡核苷酸链的合成,有望推动DNA合成技术的再次升级[2].随着DNA合成技术的快速发展,合成生物学在多个研究方向不断取得突破.本文将对基因回路、基因组设计合成、细胞工厂和人工多细胞体系的重要研究进展进行综述,并对合成生物学与其他学科交叉融合产生的一些新研究方向进行简介. ...
De novo DNA synthesis using polymerase-nucleotide conjugates
1
2018
... DNA的人工合成是合成生物学研究的底层推动技术.DNA化学合成法是当前主流的商业化合成方法,在合成长度和合成通量上不断取得突破,发展出柱式合成和芯片合成工艺,使寡核苷酸链合成通量可以达到百万条级,合成成本降低约3个数量级[1].近年来,DNA酶促合成法日益受到关注,利用末端脱氧核苷酸转移酶(TdT)在温和的条件进行寡核苷酸链的合成,有望推动DNA合成技术的再次升级[2].随着DNA合成技术的快速发展,合成生物学在多个研究方向不断取得突破.本文将对基因回路、基因组设计合成、细胞工厂和人工多细胞体系的重要研究进展进行综述,并对合成生物学与其他学科交叉融合产生的一些新研究方向进行简介. ...
A synthetic oscillatory network of transcriptional regulators
1
2000
... 基因回路是由调控元件和被调控的基因构成的特定逻辑关系,以实现所设计的预期功能.“双稳态开关”(Toggle switch)基因回路及“自激振荡网络”(Oscillatory network)基因回路的研究开启了这一合成生物学基础研究分支[3,4].基因回路往往借鉴电器回路的逻辑控制概念,与底盘细胞基因网络形成一定程度的正交关系,其未来的一个重要发展方向是构建“生物计算机”的“编程与控制”系统.基因回路的要素有:标准化的基因元件库、多基因之间的调控逻辑、调谐与降噪等回路整体的维护,最终实现由简单到复杂的目标功能(图1).由基因回路组装更大规模的网络时,对模块化封装提出需求.此外,当搭载基因回路的个体细胞以群体形式执行功能时,也需要细胞间能够通讯、互作、分工. ...
Construction of a genetic toggle switch in Escherichia coli
2
2000
... 基因回路是由调控元件和被调控的基因构成的特定逻辑关系,以实现所设计的预期功能.“双稳态开关”(Toggle switch)基因回路及“自激振荡网络”(Oscillatory network)基因回路的研究开启了这一合成生物学基础研究分支[3,4].基因回路往往借鉴电器回路的逻辑控制概念,与底盘细胞基因网络形成一定程度的正交关系,其未来的一个重要发展方向是构建“生物计算机”的“编程与控制”系统.基因回路的要素有:标准化的基因元件库、多基因之间的调控逻辑、调谐与降噪等回路整体的维护,最终实现由简单到复杂的目标功能(图1).由基因回路组装更大规模的网络时,对模块化封装提出需求.此外,当搭载基因回路的个体细胞以群体形式执行功能时,也需要细胞间能够通讯、互作、分工. ...
... 基因回路最常用的调控逻辑参考电器元件中的“开关”与“逻辑门”.基因开关包含多种形式,如能在两个基因表达稳态之间切换的双稳态开关[4]、能被特定化学小分子诱导开启的核糖开关[17]、能由RNA变构进行控制的RNA开关[18]等.对于基因逻辑门,可在单个细胞中建立转录级联控制的分层逻辑门[19],也可利用重组酶的通用特性建立16种“2输入1输出型”布尔逻辑门[20].基因开关与逻辑门的开发增强了细胞选择与执行复杂行为的能力. ...
Synthetic toolkit for complex genetic circuit engineering in Saccharomyces cerevisiae
1
2018
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
Insulated transcriptional elements enable precise design of genetic circuits
1
2017
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
Engineering BioBrick vectors from BioBrick parts
1
2008
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
BglBrick vectors and datasheets: a synthetic biology platform for gene expression
1
2011
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
Yeast golden gate (yGG) for the efficient assembly of s-cerevisiae transcription Units
1
2015
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
A modular cloning system for standardized assembly of multigene constructs
1
2011
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
A sigma factor toolbox for orthogonal gene expression in Escherichia coli
1
2018
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
The development and characterization of synthetic minimal yeast promoters
1
2015
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
Biosensor-based engineering of biosynthetic pathways
1
2016
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
Modular control of multiple pathways using engineered orthogonal T7 polymerases
1
2012
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
T7 polymerase expression of guide RNAs in vivo allows exportable CRISPR-Cas9 editing in multiple yeast hosts
1
2018
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
Functional characterization of insulation effect for synthetic gene circuits in mammalian cells
1
2018
... 基因元件是控制基因转录与翻译过程的生物分子及其对应的DNA序列,如启动子、终止子、阻遏蛋白、转录激活序列、RNA发夹结构、重组酶等.元件在回路中的逻辑作用可分为带有阈值的开启或激活逻辑以及关闭或抑制逻辑.在常用微生物模式底盘中,已实现标准化的元件库及元件之间的拼装方法[5,6].基于内切酶产生标准化的黏性末端,再由这些末端进行精确识别,在连接酶的作用下将元件拼装在一起,依据这个原理相继开发出了Biobrick[7]、Bglbrick[8]、yGG[9]、MoClo[10]等元件库及拼装技术流程.针对元件序列内部也建立了分块设计方案,用于微调基因的表达.原核启动子的Sigma转录因子、AT富含区、核糖体结合的SD序列等,以及真核启动子的转录激活位点、TATA框、转录起始位点等,均可进行标准化设计[11,12].此外,由信号分子控制的指定蛋白与操纵子位点结合[13]以及T7 RNA聚合酶[14,15]则在原核与真核细胞中都能使用,具有通用性特征.此外,在哺乳动物等高等生物中也已成功开发出调控作用元件,例如:利用3种绝缘子可对6种启动子进行交叉调节[16]. ...
Structure of a natural guanine-responsive riboswitch complexed with the metabolite hypoxanthine
1
2004
... 基因回路最常用的调控逻辑参考电器元件中的“开关”与“逻辑门”.基因开关包含多种形式,如能在两个基因表达稳态之间切换的双稳态开关[4]、能被特定化学小分子诱导开启的核糖开关[17]、能由RNA变构进行控制的RNA开关[18]等.对于基因逻辑门,可在单个细胞中建立转录级联控制的分层逻辑门[19],也可利用重组酶的通用特性建立16种“2输入1输出型”布尔逻辑门[20].基因开关与逻辑门的开发增强了细胞选择与执行复杂行为的能力. ...
Structure of an RNA switch that enforces stringent retroviral genomic RNA dimerization
1
2006
... 基因回路最常用的调控逻辑参考电器元件中的“开关”与“逻辑门”.基因开关包含多种形式,如能在两个基因表达稳态之间切换的双稳态开关[4]、能被特定化学小分子诱导开启的核糖开关[17]、能由RNA变构进行控制的RNA开关[18]等.对于基因逻辑门,可在单个细胞中建立转录级联控制的分层逻辑门[19],也可利用重组酶的通用特性建立16种“2输入1输出型”布尔逻辑门[20].基因开关与逻辑门的开发增强了细胞选择与执行复杂行为的能力. ...
Genetic programs constructed from layered logic gates in single cells
1
2012
... 基因回路最常用的调控逻辑参考电器元件中的“开关”与“逻辑门”.基因开关包含多种形式,如能在两个基因表达稳态之间切换的双稳态开关[4]、能被特定化学小分子诱导开启的核糖开关[17]、能由RNA变构进行控制的RNA开关[18]等.对于基因逻辑门,可在单个细胞中建立转录级联控制的分层逻辑门[19],也可利用重组酶的通用特性建立16种“2输入1输出型”布尔逻辑门[20].基因开关与逻辑门的开发增强了细胞选择与执行复杂行为的能力. ...
Synthetic circuits integrating logic and memory in living cells
1
2013
... 基因回路最常用的调控逻辑参考电器元件中的“开关”与“逻辑门”.基因开关包含多种形式,如能在两个基因表达稳态之间切换的双稳态开关[4]、能被特定化学小分子诱导开启的核糖开关[17]、能由RNA变构进行控制的RNA开关[18]等.对于基因逻辑门,可在单个细胞中建立转录级联控制的分层逻辑门[19],也可利用重组酶的通用特性建立16种“2输入1输出型”布尔逻辑门[20].基因开关与逻辑门的开发增强了细胞选择与执行复杂行为的能力. ...
A fast, robust and tunable synthetic gene oscillator
1
2008
... 基因回路需要对多个基因表达进行精细控制,才能实现整体的精细功能.利用降解标签来控制元件的蛋白质降解时间,可以设计构建出频率与振幅均可自由调控的振荡器装置[21].利用逻辑“非门”检查细胞对时序逻辑的执行,从而达到回路时序控制[22].通过构建“长寿命基因时钟”,可延长基因回路在细胞中发挥功能的时间,实现回路的时限控制[23].通过调控元件生物分子的降解速率,可将振荡器振荡周期的标准差从35%降低到14%,达到回路降噪的目的[24]. ...
Cellular checkpoint control using programmable sequential logic
1
2018
... 基因回路需要对多个基因表达进行精细控制,才能实现整体的精细功能.利用降解标签来控制元件的蛋白质降解时间,可以设计构建出频率与振幅均可自由调控的振荡器装置[21].利用逻辑“非门”检查细胞对时序逻辑的执行,从而达到回路时序控制[22].通过构建“长寿命基因时钟”,可延长基因回路在细胞中发挥功能的时间,实现回路的时限控制[23].通过调控元件生物分子的降解速率,可将振荡器振荡周期的标准差从35%降低到14%,达到回路降噪的目的[24]. ...
Rock-paper-scissors: engineered population dynamics increase genetic stability
1
2019
... 基因回路需要对多个基因表达进行精细控制,才能实现整体的精细功能.利用降解标签来控制元件的蛋白质降解时间,可以设计构建出频率与振幅均可自由调控的振荡器装置[21].利用逻辑“非门”检查细胞对时序逻辑的执行,从而达到回路时序控制[22].通过构建“长寿命基因时钟”,可延长基因回路在细胞中发挥功能的时间,实现回路的时限控制[23].通过调控元件生物分子的降解速率,可将振荡器振荡周期的标准差从35%降低到14%,达到回路降噪的目的[24]. ...
Synchronous long-term oscillations in a synthetic gene circuit
1
2016
... 基因回路需要对多个基因表达进行精细控制,才能实现整体的精细功能.利用降解标签来控制元件的蛋白质降解时间,可以设计构建出频率与振幅均可自由调控的振荡器装置[21].利用逻辑“非门”检查细胞对时序逻辑的执行,从而达到回路时序控制[22].通过构建“长寿命基因时钟”,可延长基因回路在细胞中发挥功能的时间,实现回路的时限控制[23].通过调控元件生物分子的降解速率,可将振荡器振荡周期的标准差从35%降低到14%,达到回路降噪的目的[24]. ...
Programmable protein circuits in living cells
1
2018
... 当不同的基因回路组成更加复杂的基因调控网络时,需要对回路进行一定程度的模块化封装.利用正交化-模块化病毒蛋白酶可对靶蛋白进行对接与切割,实现了8种二进制逻辑门和动态模拟信号处理等封装功能[25].将基因回路包裹在脂质体小滴中,添加基因表达所需工具酶等组分,可控制各回路独立工作与产物交流[26].为了对基因回路的设计进行加密,可分别建立“加密回路拓扑”与“模糊化回路连接”策略,使得未经授权则难以确定回路结构和功能[27]. ...
Engineering genetic circuit interactions within and between synthetic minimal cells
1
2017
... 当不同的基因回路组成更加复杂的基因调控网络时,需要对回路进行一定程度的模块化封装.利用正交化-模块化病毒蛋白酶可对靶蛋白进行对接与切割,实现了8种二进制逻辑门和动态模拟信号处理等封装功能[25].将基因回路包裹在脂质体小滴中,添加基因表达所需工具酶等组分,可控制各回路独立工作与产物交流[26].为了对基因回路的设计进行加密,可分别建立“加密回路拓扑”与“模糊化回路连接”策略,使得未经授权则难以确定回路结构和功能[27]. ...
Encryption and steganography of synthetic gene circuits
1
2018
... 当不同的基因回路组成更加复杂的基因调控网络时,需要对回路进行一定程度的模块化封装.利用正交化-模块化病毒蛋白酶可对靶蛋白进行对接与切割,实现了8种二进制逻辑门和动态模拟信号处理等封装功能[25].将基因回路包裹在脂质体小滴中,添加基因表达所需工具酶等组分,可控制各回路独立工作与产物交流[26].为了对基因回路的设计进行加密,可分别建立“加密回路拓扑”与“模糊化回路连接”策略,使得未经授权则难以确定回路结构和功能[27]. ...
Artificial cell-cell communication in yeast Saccharomyces cerevisiae using signaling elements from Arabidopsis thaliana
1
2005
... 搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系.“群体感应”基因回路已从原核细胞拓展到真核细胞中[28,29].基因回路控制的亚细胞群可形成“捕食者”关系模型[30]、细胞“荧光条纹分布”模型[31]等.通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门[32].通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算[33].个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构[34]. ...
Programmed population control by cell-cell communication and regulated killing
1
2004
... 搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系.“群体感应”基因回路已从原核细胞拓展到真核细胞中[28,29].基因回路控制的亚细胞群可形成“捕食者”关系模型[30]、细胞“荧光条纹分布”模型[31]等.通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门[32].通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算[33].个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构[34]. ...
A synthetic Escherichia coli predator-prey ecosystem
2
2008
... 搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系.“群体感应”基因回路已从原核细胞拓展到真核细胞中[28,29].基因回路控制的亚细胞群可形成“捕食者”关系模型[30]、细胞“荧光条纹分布”模型[31]等.通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门[32].通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算[33].个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构[34]. ...
... 在互利共生体系中,菌株之间共享或者交换营养物质,彼此之间可共同完成某一种化合物的彻底代谢,并分别从中获益.通过构建了营养缺陷型菌株,可以构建互利共生的多细胞系统,使菌株促进彼此的生长[165].研究者利用双向通讯在两株大肠杆菌中构建了共生系统[166].在偏利或偏害关系的体系中,对一方菌株有益/有害,另一方则不受影响.菌株间对环境资源(如营养物质、空间等)的争夺则形成了菌株间的竞争关系.在构建人工多细胞体系时,也应充分考虑菌株间竞争、偏利或偏害等关系存在的可能性.部分菌株可能加速对营养物质的吸收,并对这些物质进行与构建目标不相关的无效转化,或者产生对其他菌株有抑制作用的物质,这必然会影响菌群的高效和稳定.通过对混菌体系中菌株关系的重构,可解除竞争抑制,使菌株间偏利共生和竞争关系转变为互利共生[167].通过人工构建和调控菌株间的互利共生关系,使人工多细胞体系对复杂功能网络进行有效的分工协作,可实现混菌系统的高效、稳定、可控.此外,研究者还设计构建了多种菌群相互作用关系,为构建人工多细胞体系提供新的思路.例如:在两株大肠杆菌中分别构建群体感应信号调控下的细胞自杀基因表达及其抑制途径,可形成捕食者与被捕食者的关系[30]. ...
Sequential establishment of stripe patterns in an expanding cell population
1
2011
... 搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系.“群体感应”基因回路已从原核细胞拓展到真核细胞中[28,29].基因回路控制的亚细胞群可形成“捕食者”关系模型[30]、细胞“荧光条纹分布”模型[31]等.通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门[32].通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算[33].个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构[34]. ...
Robust multicellular computing using genetically encoded NOR gates and chemical 'wires'
1
2011
... 搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系.“群体感应”基因回路已从原核细胞拓展到真核细胞中[28,29].基因回路控制的亚细胞群可形成“捕食者”关系模型[30]、细胞“荧光条纹分布”模型[31]等.通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门[32].通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算[33].个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构[34]. ...
Distributed biological computation with multicellular engineered networks
1
2011
... 搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系.“群体感应”基因回路已从原核细胞拓展到真核细胞中[28,29].基因回路控制的亚细胞群可形成“捕食者”关系模型[30]、细胞“荧光条纹分布”模型[31]等.通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门[32].通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算[33].个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构[34]. ...
Programming self-organizing multicellular structures with synthetic cell-cell signaling
1
2018
... 搭载回路的多个细胞组成的细胞群可构成协同、竞争等不同的群体关系.“群体感应”基因回路已从原核细胞拓展到真核细胞中[28,29].基因回路控制的亚细胞群可形成“捕食者”关系模型[30]、细胞“荧光条纹分布”模型[31]等.通过建立大肠杆菌菌斑之间的通讯,可构建所有双输入布尔逻辑门[32].通过混养搭载不同基因回路的酵母细胞亚群,可实现多路选择器和进位加法器的逻辑运算[33].个体细胞之间也可发展成可控的多细胞结构,例如通过建立动物细胞模块化分泌信号平台,可控制细胞表面钙黏蛋白的表达,从而控制多细胞间的黏附结合与整体结构[34]. ...
Design of a dynamic sensor-regulator system for production of chemicals and fuels derived from fatty acids
2
2012
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Programming adaptive control to evolve increased metabolite production
1
2013
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
Genetically encoded fluorescent sensors reveal dynamic regulation of NADPH metabolism
1
2017
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
A Synthetic circuit for mercury bioremediation using self assembling functional amyloids
1
2017
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
Cascaded amplifying circuits enable ultrasensitive cellular sensors for toxic metals
1
2019
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
Multilayered genetic safeguards limit growth of microorganisms to defined environments
1
2015
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
'Deadman' and 'Passcode' microbial kill switches for bacterial containment
1
2016
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
Precision tumor recognition by T cells with combinatorial antigen-sensing circuits
1
2016
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
Engineering T cells with customized therapeutic response programs using synthetic notch receptors
1
2016
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
An ingestible bacterial-electronic system to monitor gastrointestinal health
1
2018
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
Smartphone-controlled optogenetically engineered cells enable semiautomatic glucose homeostasis in diabetic mice
1
2017
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
A synthetic low-frequency mammalian oscillator
1
2010
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
A tunable synthetic mammalian oscillator
1
2009
... 近年来,基因回路逐步走向生物制造、环境监测、生物安防、医疗等应用领域.最常使用的是成套的“传感器+开关/逻辑门”回路,通过感应特定变化,细胞将开启或关闭指定功能,或在不同功能间切换.研究者已开发了响应细胞内特殊代谢物(如脂肪酸[35]、异戊二烯[36])、响应辅酶及辅因子[37]等的传感器开关,用于大幅提升目标生物制品的产量.基因回路可用于砷、汞等环境毒性物质的检测,提升其检测灵敏度,降低环境有害污染物的监测成本[38,39].越来越多的携带基因回路的微生物被实际应用,防逃逸回路的开发非常必要.可利用外加小分子控制核糖开关和核酸酶元件,控制大肠杆菌的生存[40],也可设计特定的“死亡回路”和“密码”来控制细菌的死亡[41].在医疗领域,基因回路在嵌合抗原受体T细胞(CAR-T)治疗中发挥了重要作用.由鼠Notch跨膜受体改造而成的工程化SynNotch受体,可使T细胞只有先后识别两种特异性抗原时才被激活;基于SynNotch的逻辑“与门”,也可利用肿瘤抗原和特定小分子药物作为输入端,有效降低IL-12等细胞因子风暴[42,43].通过设计可摄入的微生物电子设备,可对生物分子进行原位检测,例如利用血红素敏感的益生菌传感器诊断猪胃肠道出血的发生[44].传感型基因回路在糖尿病治疗中也可发挥作用,例如:利用智能手机远程控制小鼠中特定基因表达,以及用咖啡因小分子诱导控制小鼠细胞生成治疗Ⅱ型糖尿病的多肽,从而调节血糖水平[45].此外,利用哺乳动物细胞振荡器回路控制基因周期性表达也逐步走向治疗应用[46,47]. ...
Creation of a bacterial cell controlled by a chemically synthesized genome
2
2010
... 2010年,Synthia细胞的自我复制标志着人工合成基因组实现了对生命活动的调控,突破了化学物质和活性基因组的界限[48].人工基因组合成是关于理性设计和重新合成生命的研究,即在工程学思想的指导下,借助计算机模拟,模块化设计具有特定功能的人工基因组,利用DNA从头合成和模块化组装技术,将人工设计基因组构建出来,并使其实现预期功能. ...
... 人工基因组的长度远远超出了普通DNA化学合成的长度范围,需要通过标准化DNA元件的逐级组装技术来实现.根据组装原理不同,可将DNA元件组装技术分为酶促体外组装(基于DNA聚合酶、核酸内切酶或核酸外切酶)、非酶促体外组装和体内组装(基于DNA同源重组或位点特异性重组).酶促体外组装和体内组装技术已应用于支原体、大肠杆菌和酿酒酵母的人工基因组合成,推动了组装大片段DNA(kb至Mb级)的发展[48,51,52,53]. ...
Design and synthesis of a minimal bacterial genome
2
2016
... 近年来,人工基因组合成取得了一系列重大突破.最小化基因组的理性合成颠覆了简化生命体的传统策略,使我们对人工细胞在特定环境下的行为和功能机制的理解更加深入[49].密码子转换和非天然氨基酸技术的应用实现了正交化生命体的创建,拓展了生命进化方向和生命存在形式的可能性[50,51].人工基因组合成的发展内容主要包括DNA片段高效组装和迭代替换、基因组精简与遗传密码扩展、遗传系统可控进化等,最终实现人工细胞性能的定向优化(图2). ...
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
Design, synthesis, and testing toward a 57-codon genome
3
2016
... 近年来,人工基因组合成取得了一系列重大突破.最小化基因组的理性合成颠覆了简化生命体的传统策略,使我们对人工细胞在特定环境下的行为和功能机制的理解更加深入[49].密码子转换和非天然氨基酸技术的应用实现了正交化生命体的创建,拓展了生命进化方向和生命存在形式的可能性[50,51].人工基因组合成的发展内容主要包括DNA片段高效组装和迭代替换、基因组精简与遗传密码扩展、遗传系统可控进化等,最终实现人工细胞性能的定向优化(图2). ...
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
... [50,51,53,54,55,56,57,58]. ...
Total synthesis of Escherichia coli with a recoded genome
7
2019
... 近年来,人工基因组合成取得了一系列重大突破.最小化基因组的理性合成颠覆了简化生命体的传统策略,使我们对人工细胞在特定环境下的行为和功能机制的理解更加深入[49].密码子转换和非天然氨基酸技术的应用实现了正交化生命体的创建,拓展了生命进化方向和生命存在形式的可能性[50,51].人工基因组合成的发展内容主要包括DNA片段高效组装和迭代替换、基因组精简与遗传密码扩展、遗传系统可控进化等,最终实现人工细胞性能的定向优化(图2). ...
... 人工基因组的长度远远超出了普通DNA化学合成的长度范围,需要通过标准化DNA元件的逐级组装技术来实现.根据组装原理不同,可将DNA元件组装技术分为酶促体外组装(基于DNA聚合酶、核酸内切酶或核酸外切酶)、非酶促体外组装和体内组装(基于DNA同源重组或位点特异性重组).酶促体外组装和体内组装技术已应用于支原体、大肠杆菌和酿酒酵母的人工基因组合成,推动了组装大片段DNA(kb至Mb级)的发展[48,51,52,53]. ...
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
... ,51,53,54,55,56,57,58]. ...
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
... 人工基因组合成研究显著提升了获得基因组变异的能力,可以为人工细胞进化和演化提供驱动力,为深入探索系统进化和物种演化的分子机制提供了新的平台.人工基因组的进化和演化在原核和真核细胞中开展,主要包括重叠基因的重构、基因组诱导重排等.人工大肠杆菌基因组中79个重叠基因的重构,实现了基因重叠区域的独立转录[51],探究了基因编码元件交替分布排列的演化特征.人工酵母基因组的诱导重排研究,揭示了特殊环境下的基因组变异规律和适应性演化机制;并利用该技术开展高产植物源化合物的人工酵母底盘细胞构建和功能基因组研究,提升胡萝卜素产量38.8倍,发现决定重要性状的关键基因,阐明基因表达调控、代谢调节等网络的组成和变化[53,64,65]. ...
... 遗传密码拓展技术是非天然氨基酸引入的重要手段,主要是通过控制终止密码子的读取,进而通过氨酰-tRNA合成酶/tRNA对的改造实现[217].近年来基因组密码子删减研究可以为遗传密码拓展提供高效的人工底盘细胞[51,60],将明显提升引入非天然氨基酸的数量、种类和效率.此外,非天然核酸加入基因组的研究也将为更多非天然氨基酸的进一步高效加入提供新的方向[218]. ...
Total synthesis of a functional designer eukaryotic chromosome
1
2014
... 人工基因组的长度远远超出了普通DNA化学合成的长度范围,需要通过标准化DNA元件的逐级组装技术来实现.根据组装原理不同,可将DNA元件组装技术分为酶促体外组装(基于DNA聚合酶、核酸内切酶或核酸外切酶)、非酶促体外组装和体内组装(基于DNA同源重组或位点特异性重组).酶促体外组装和体内组装技术已应用于支原体、大肠杆菌和酿酒酵母的人工基因组合成,推动了组装大片段DNA(kb至Mb级)的发展[48,51,52,53]. ...
Perfect designer chromosome Ⅴ and behavior of a ring derivative
3
2017
... 人工基因组的长度远远超出了普通DNA化学合成的长度范围,需要通过标准化DNA元件的逐级组装技术来实现.根据组装原理不同,可将DNA元件组装技术分为酶促体外组装(基于DNA聚合酶、核酸内切酶或核酸外切酶)、非酶促体外组装和体内组装(基于DNA同源重组或位点特异性重组).酶促体外组装和体内组装技术已应用于支原体、大肠杆菌和酿酒酵母的人工基因组合成,推动了组装大片段DNA(kb至Mb级)的发展[48,51,52,53]. ...
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
... 人工基因组合成研究显著提升了获得基因组变异的能力,可以为人工细胞进化和演化提供驱动力,为深入探索系统进化和物种演化的分子机制提供了新的平台.人工基因组的进化和演化在原核和真核细胞中开展,主要包括重叠基因的重构、基因组诱导重排等.人工大肠杆菌基因组中79个重叠基因的重构,实现了基因重叠区域的独立转录[51],探究了基因编码元件交替分布排列的演化特征.人工酵母基因组的诱导重排研究,揭示了特殊环境下的基因组变异规律和适应性演化机制;并利用该技术开展高产植物源化合物的人工酵母底盘细胞构建和功能基因组研究,提升胡萝卜素产量38.8倍,发现决定重要性状的关键基因,阐明基因表达调控、代谢调节等网络的组成和变化[53,64,65]. ...
Design of a synthetic yeast genome
2
2017
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
Synthesis, debugging, and effects of synthetic chromosome consolidation: syn Ⅵ and beyond
1
2017
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
Deep functional analysis of synII, a 770-kilobase synthetic yeast chromosome
1
2017
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
Bug mapping and fitness testing of chemically synthesized chromosome X
1
2017
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
Engineering the ribosomal DNA in a megabase synthetic chromosome
1
2017
... 随后,标准化DNA元件在细胞内对天然基因组进行迭代替换,直至获得完整人工基因组.位点特异性重组和同源重组技术分别应用于原核和真核基因组的迭代替换,同时基因编辑技术大幅提高DNA迭代替换的效率[50,51].为进一步加速人工基因组合成的速度,不同的DNA元件分别在多个细胞中进行迭代替换,通过细胞融合进行基因组的转移,快速获得具有完整活性的人工基因组[50,51,53,54,55,56,57,58]. ...
Design and chemical synthesis of eukaryotic chromosomes
1
2017
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
Precise manipulation of chromosomes in vivo enables genome-wide codon replacement
2
2011
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
... 遗传密码拓展技术是非天然氨基酸引入的重要手段,主要是通过控制终止密码子的读取,进而通过氨酰-tRNA合成酶/tRNA对的改造实现[217].近年来基因组密码子删减研究可以为遗传密码拓展提供高效的人工底盘细胞[51,60],将明显提升引入非天然氨基酸的数量、种类和效率.此外,非天然核酸加入基因组的研究也将为更多非天然氨基酸的进一步高效加入提供新的方向[218]. ...
Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage
1
2016
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
Programmable base editing of A?T to G?C in genomic DNA without DNA cleavage
1
2017
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
Genome-scale engineering of Saccharomyces cerevisiae with single-nucleotide precision
1
2018
... 人工基因组合成能力的提升推动了基因组设计深度的扩展,可以进行复杂遗传信息的精简,重塑细胞生命活动.基因组精简的研究在支原体、大肠杆菌和酿酒酵母等多个物种中展开,主要策略包括非必需基因精简、遗传元件的移除或移位、密码子的简化等.在最小支原体基因组的构建中,精简了大量非必需基因和半必需基因,获得的最小支原体基因组长度缩减50%,仅由473个基因组成[49].在酿酒酵母人工基因组Sc2.0的设计中,删除了转座序列、内含子、亚端粒等遗传元件,对tRNA基因进行重新定位,人工酵母基因组序列精简了6%[54,59].基因组合成可以实现全基因组序列的密码子转换,已实现大肠杆菌基因组1.8万个靶标密码子的转换,精简了丝氨酸密码子TCG、TCA和终止密码子TAG,构建了仅有61个遗传密码的人工大肠杆菌[51].基因编辑以天然的生物基因组为蓝本,可以修饰和改造特定位点的DNA序列,已实现对大肠杆菌基因组314个TAG终止密码子的转换[60].随着基因编辑效率的提高、脱靶效应的降低和新方法的不断更新,尤其是碱基编辑器技术的发展,可进行密码子转换的基因组DNA序列区域不断扩大,基因编辑将推动基因组设计与合成的快速发展[61,62,63]. ...
Precise control of SCRaMbLE in synthetic haploid and diploid yeast
1
2018
... 人工基因组合成研究显著提升了获得基因组变异的能力,可以为人工细胞进化和演化提供驱动力,为深入探索系统进化和物种演化的分子机制提供了新的平台.人工基因组的进化和演化在原核和真核细胞中开展,主要包括重叠基因的重构、基因组诱导重排等.人工大肠杆菌基因组中79个重叠基因的重构,实现了基因重叠区域的独立转录[51],探究了基因编码元件交替分布排列的演化特征.人工酵母基因组的诱导重排研究,揭示了特殊环境下的基因组变异规律和适应性演化机制;并利用该技术开展高产植物源化合物的人工酵母底盘细胞构建和功能基因组研究,提升胡萝卜素产量38.8倍,发现决定重要性状的关键基因,阐明基因表达调控、代谢调节等网络的组成和变化[53,64,65]. ...
Ring synthetic chromosome ⅤSCRaMbLE
1
2018
... 人工基因组合成研究显著提升了获得基因组变异的能力,可以为人工细胞进化和演化提供驱动力,为深入探索系统进化和物种演化的分子机制提供了新的平台.人工基因组的进化和演化在原核和真核细胞中开展,主要包括重叠基因的重构、基因组诱导重排等.人工大肠杆菌基因组中79个重叠基因的重构,实现了基因重叠区域的独立转录[51],探究了基因编码元件交替分布排列的演化特征.人工酵母基因组的诱导重排研究,揭示了特殊环境下的基因组变异规律和适应性演化机制;并利用该技术开展高产植物源化合物的人工酵母底盘细胞构建和功能基因组研究,提升胡萝卜素产量38.8倍,发现决定重要性状的关键基因,阐明基因表达调控、代谢调节等网络的组成和变化[53,64,65]. ...
Metabolic engineering for the microbial production of 1,3-propanediol
1
2003
... 构建细胞工厂从头合成生物基材料单体1,3-丙二醇[66]和重要医药中间体青蒿酸[67,68],开创了传统石化产品和天然产物全新的生产模式,可有效解决石油化工炼制和植物提取对自然资源的依赖和对环境的危害.自此,细胞工厂的设计构建成为合成生物学的一个重要研究方向.伴随着合成生物技术的进步,人们在逐步挑战代谢途径更长、复杂程度更高的化合物合成[图3(a)].尤其是近期阿片类药物[69]和大麻素[70]的从头合成,展示了构建细胞工厂的巨大潜力.细胞工厂的设计构建,是通过对复杂生命体的工程化重构,实现目标产品的可控和高效合成.其实现过程是在物质代谢和能量代谢水平上,反复进行模块与模块、模块与底盘之间的适配[71].除适配性外,细胞工厂的发展趋势还承袭了模块化、正交性、鲁棒性等工程化特性. ...
Production of the antimalarial drug precursor artemisinic acid in engineered yeast
1
2006
... 构建细胞工厂从头合成生物基材料单体1,3-丙二醇[66]和重要医药中间体青蒿酸[67,68],开创了传统石化产品和天然产物全新的生产模式,可有效解决石油化工炼制和植物提取对自然资源的依赖和对环境的危害.自此,细胞工厂的设计构建成为合成生物学的一个重要研究方向.伴随着合成生物技术的进步,人们在逐步挑战代谢途径更长、复杂程度更高的化合物合成[图3(a)].尤其是近期阿片类药物[69]和大麻素[70]的从头合成,展示了构建细胞工厂的巨大潜力.细胞工厂的设计构建,是通过对复杂生命体的工程化重构,实现目标产品的可控和高效合成.其实现过程是在物质代谢和能量代谢水平上,反复进行模块与模块、模块与底盘之间的适配[71].除适配性外,细胞工厂的发展趋势还承袭了模块化、正交性、鲁棒性等工程化特性. ...
Engineering a mevalonate pathway in Escherichia coli for production of terpenoids
1
2003
... 构建细胞工厂从头合成生物基材料单体1,3-丙二醇[66]和重要医药中间体青蒿酸[67,68],开创了传统石化产品和天然产物全新的生产模式,可有效解决石油化工炼制和植物提取对自然资源的依赖和对环境的危害.自此,细胞工厂的设计构建成为合成生物学的一个重要研究方向.伴随着合成生物技术的进步,人们在逐步挑战代谢途径更长、复杂程度更高的化合物合成[图3(a)].尤其是近期阿片类药物[69]和大麻素[70]的从头合成,展示了构建细胞工厂的巨大潜力.细胞工厂的设计构建,是通过对复杂生命体的工程化重构,实现目标产品的可控和高效合成.其实现过程是在物质代谢和能量代谢水平上,反复进行模块与模块、模块与底盘之间的适配[71].除适配性外,细胞工厂的发展趋势还承袭了模块化、正交性、鲁棒性等工程化特性. ...
Total biosynthesis of opiates by stepwise fermentation using engineered Escherichia coli
2
2016
... 构建细胞工厂从头合成生物基材料单体1,3-丙二醇[66]和重要医药中间体青蒿酸[67,68],开创了传统石化产品和天然产物全新的生产模式,可有效解决石油化工炼制和植物提取对自然资源的依赖和对环境的危害.自此,细胞工厂的设计构建成为合成生物学的一个重要研究方向.伴随着合成生物技术的进步,人们在逐步挑战代谢途径更长、复杂程度更高的化合物合成[图3(a)].尤其是近期阿片类药物[69]和大麻素[70]的从头合成,展示了构建细胞工厂的巨大潜力.细胞工厂的设计构建,是通过对复杂生命体的工程化重构,实现目标产品的可控和高效合成.其实现过程是在物质代谢和能量代谢水平上,反复进行模块与模块、模块与底盘之间的适配[71].除适配性外,细胞工厂的发展趋势还承袭了模块化、正交性、鲁棒性等工程化特性. ...
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Complete biosynthesis of cannabinoids and their unnatural analogues in yeast
1
2019
... 构建细胞工厂从头合成生物基材料单体1,3-丙二醇[66]和重要医药中间体青蒿酸[67,68],开创了传统石化产品和天然产物全新的生产模式,可有效解决石油化工炼制和植物提取对自然资源的依赖和对环境的危害.自此,细胞工厂的设计构建成为合成生物学的一个重要研究方向.伴随着合成生物技术的进步,人们在逐步挑战代谢途径更长、复杂程度更高的化合物合成[图3(a)].尤其是近期阿片类药物[69]和大麻素[70]的从头合成,展示了构建细胞工厂的巨大潜力.细胞工厂的设计构建,是通过对复杂生命体的工程化重构,实现目标产品的可控和高效合成.其实现过程是在物质代谢和能量代谢水平上,反复进行模块与模块、模块与底盘之间的适配[71].除适配性外,细胞工厂的发展趋势还承袭了模块化、正交性、鲁棒性等工程化特性. ...
DCEO biotechnology: tools to design, construct, evaluate, and optimize the metabolic pathway for biosynthesis of chemicals
1
2018
... 构建细胞工厂从头合成生物基材料单体1,3-丙二醇[66]和重要医药中间体青蒿酸[67,68],开创了传统石化产品和天然产物全新的生产模式,可有效解决石油化工炼制和植物提取对自然资源的依赖和对环境的危害.自此,细胞工厂的设计构建成为合成生物学的一个重要研究方向.伴随着合成生物技术的进步,人们在逐步挑战代谢途径更长、复杂程度更高的化合物合成[图3(a)].尤其是近期阿片类药物[69]和大麻素[70]的从头合成,展示了构建细胞工厂的巨大潜力.细胞工厂的设计构建,是通过对复杂生命体的工程化重构,实现目标产品的可控和高效合成.其实现过程是在物质代谢和能量代谢水平上,反复进行模块与模块、模块与底盘之间的适配[71].除适配性外,细胞工厂的发展趋势还承袭了模块化、正交性、鲁棒性等工程化特性. ...
In vitro reconstitution of indolmycin biosynthesis reveals the molecular basis of oxazolinone assembly
1
2015
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Activation and characterization of a cryptic polycyclic tetramate macrolactam biosynthetic gene cluster
1
2013
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
GenomGenome of Tripterygium wilfordii and identification of cytochrome P450 involved in triptolide biosynthesis
1
2020
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Six enzymes from mayapple that complete the biosynthetic pathway to the etoposide aglycone
1
2015
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Design and evolution of enzymes for non-natural chemistry
1
2017
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Engineering new catalytic activities in enzymes
1
2020
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
ROSETTA3: an object-oriented software suite for the simulation and design of macromolecules
1
2011
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
De novo design of a fluorescence-activating beta-barrel
1
2018
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
De novo design of tunable, pH-driven conformational changes
1
2019
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Programmable design of orthogonal protein heterodimers
1
2019
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Anti-Markovnikov alkene oxidation by metal-oxo-mediated enzyme catalysis
1
2017
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Genetically programmed chiral organoborane synthesis
1
2017
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Directed evolution of cytochrome c for carbon-silicon bond formation: bringing silicon to life
1
2016
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Synthesis of 11-carbon terpenoids in yeast using protein and metabolic engineering
1
2018
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
A highly selective biosynthetic pathway to non-natural C50 carotenoids assembled from moderately selective enzymes
1
2015
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Scalable production of mechanically tunable block polymers from sugar
1
2014
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Engineering nonphosphorylative metabolism to generate lignocellulose-derived products
1
2016
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Metabolic engineering of Escherichia coli for production of salvianic acid A via an artificial biosynthetic pathway
1
2013
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli
3
2010
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
... [90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
... [90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Modular optimization of multi-gene pathways for fatty acids production in E. coli
1
2013
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Modular pathway rewiring of Saccharomyces cerevisiae enables high-level production of L-ornithine
1
2015
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals
1
2016
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Metabolic engineering of Escherichia coli for de novo biosynthesis of vitamin B12
1
2018
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
Engineering a microbial biosynthesis platform for de novo production of tropane alkaloids
1
2019
... 代谢途径的模块化是细胞工厂构建的基础.模块中的催化元件或来源于对沉默基因簇[72,73]和深度测序数据[74,75]的挖掘,或来源于现有酶分子的定向进化[76,77]甚至蛋白质的从头设计[78,79,80,81],以实现自然界中不能催化或难催化的反应[82,83,84],合成非天然的分子[85,86,87],构建未见报道的合成路径[88,89].在重构复杂长途径时,采用多元模块工程[90],按代谢节点[90]和元件功能聚类[91]等规则对途径进行模块式划分[图3(b)],以降低途径的复杂程度.通过建立并优化模块功能[92]、调整模块亚细胞器定位[93]以及调节模块之间的表达强度[90],实现模块与模块之间的组合设计和迭代适配.在上述模块化工程原则的指导下,成功实现阿片类药物[69]、维生素B12[94]、托品烷生物碱[95]等长途径的构建. ...
The role of dynamic enzyme assemblies and substrate channelling in metabolic regulation
1
2018
... 以途径模块作为基本调控单元,通过调整模块间表达强度,并结合底物通道作用[96]和区室化作用[97],可以平衡途径上下游或竞争性模块间的代谢流通量,避免代谢中间体和副产物的积累.对模块的表达强度调节,除进行转录水平的调控外,还可进行翻译及翻译后水平的调控.例如在酿酒酵母中通过C-PEST介导的内质网相关降解[98]和N-degron介导的泛素化降解[99]调控蛋白的半衰期,以调整代谢流分配[100,101]. ...
Harnessing yeast organelles for metabolic engineering
2
2017
... 以途径模块作为基本调控单元,通过调整模块间表达强度,并结合底物通道作用[96]和区室化作用[97],可以平衡途径上下游或竞争性模块间的代谢流通量,避免代谢中间体和副产物的积累.对模块的表达强度调节,除进行转录水平的调控外,还可进行翻译及翻译后水平的调控.例如在酿酒酵母中通过C-PEST介导的内质网相关降解[98]和N-degron介导的泛素化降解[99]调控蛋白的半衰期,以调整代谢流分配[100,101]. ...
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Protein quality control of the endoplasmic reticulum and ubiquitin-proteasome-triggered degradation of aberrant proteins: yeast pioneers the path
1
2018
... 以途径模块作为基本调控单元,通过调整模块间表达强度,并结合底物通道作用[96]和区室化作用[97],可以平衡途径上下游或竞争性模块间的代谢流通量,避免代谢中间体和副产物的积累.对模块的表达强度调节,除进行转录水平的调控外,还可进行翻译及翻译后水平的调控.例如在酿酒酵母中通过C-PEST介导的内质网相关降解[98]和N-degron介导的泛素化降解[99]调控蛋白的半衰期,以调整代谢流分配[100,101]. ...
An N-end rule pathway that recognizes proline and destroys gluconeogenic enzymes
1
2017
... 以途径模块作为基本调控单元,通过调整模块间表达强度,并结合底物通道作用[96]和区室化作用[97],可以平衡途径上下游或竞争性模块间的代谢流通量,避免代谢中间体和副产物的积累.对模块的表达强度调节,除进行转录水平的调控外,还可进行翻译及翻译后水平的调控.例如在酿酒酵母中通过C-PEST介导的内质网相关降解[98]和N-degron介导的泛素化降解[99]调控蛋白的半衰期,以调整代谢流分配[100,101]. ...
Engineered protein degradation of farnesyl pyrophosphate synthase is an effective regulatory mechanism to increase monoterpene production in Saccharomyces cerevisiae
1
2018
... 以途径模块作为基本调控单元,通过调整模块间表达强度,并结合底物通道作用[96]和区室化作用[97],可以平衡途径上下游或竞争性模块间的代谢流通量,避免代谢中间体和副产物的积累.对模块的表达强度调节,除进行转录水平的调控外,还可进行翻译及翻译后水平的调控.例如在酿酒酵母中通过C-PEST介导的内质网相关降解[98]和N-degron介导的泛素化降解[99]调控蛋白的半衰期,以调整代谢流分配[100,101]. ...
A squalene synthase protein degradation method for improved sesquiterpene production in Saccharomyces cerevisiae
1
2017
... 以途径模块作为基本调控单元,通过调整模块间表达强度,并结合底物通道作用[96]和区室化作用[97],可以平衡途径上下游或竞争性模块间的代谢流通量,避免代谢中间体和副产物的积累.对模块的表达强度调节,除进行转录水平的调控外,还可进行翻译及翻译后水平的调控.例如在酿酒酵母中通过C-PEST介导的内质网相关降解[98]和N-degron介导的泛素化降解[99]调控蛋白的半衰期,以调整代谢流分配[100,101]. ...
Engineering customized cell sensing and response behaviors using synthetic notch receptors
1
2016
... 细胞工厂的正交化设计是指对生产菌株的工程化改造,不干扰底盘原有的代谢和调控网络,以减少人工设计的复杂性.最初正交化设计主要针对导入的异源人工元器件,例如转录因子[102]、核糖体开关[103]和Cas9蛋白[104]等,以实现底盘的基因编辑和表达调控.目前正交化的概念随细胞工厂自下而上的构建过程顺应延伸,逐步扩展到对代谢途径和真核底盘细胞器的设计中. ...
Systematic evaluation of genetic and environmental factors affecting performance of translational riboswitches
1
2019
... 细胞工厂的正交化设计是指对生产菌株的工程化改造,不干扰底盘原有的代谢和调控网络,以减少人工设计的复杂性.最初正交化设计主要针对导入的异源人工元器件,例如转录因子[102]、核糖体开关[103]和Cas9蛋白[104]等,以实现底盘的基因编辑和表达调控.目前正交化的概念随细胞工厂自下而上的构建过程顺应延伸,逐步扩展到对代谢途径和真核底盘细胞器的设计中. ...
Orthogonal gene knockout and activation with a catalytically active Cas9 nuclease
1
2015
... 细胞工厂的正交化设计是指对生产菌株的工程化改造,不干扰底盘原有的代谢和调控网络,以减少人工设计的复杂性.最初正交化设计主要针对导入的异源人工元器件,例如转录因子[102]、核糖体开关[103]和Cas9蛋白[104]等,以实现底盘的基因编辑和表达调控.目前正交化的概念随细胞工厂自下而上的构建过程顺应延伸,逐步扩展到对代谢途径和真核底盘细胞器的设计中. ...
Redesigning metabolism based on orthogonality principles
3
2017
... 在物质代谢水平,代谢途径的相对正交化是指底物同化途径或产物合成途径与底盘内源代谢网络最多只以唯一的结点相连,以形成独立的模块,便于后续调控其代谢流通量[105,106].此种途径的正交化过程可视作对底盘生长和产物合成的解耦[105].在碳源利用方面,常设计构建非天然途径,或利用葡萄糖、油脂等天然碳源[图3(c),正交化途径1],或转化半纤维素水解产物(木糖、阿拉伯糖、半乳糖)等替代碳源[图3(c),正交化途径2],合成关键前体化合物.例如,合成1,4-丁二醇[107]和琥珀酸[105]的正交化途径.在前体供给方面,常基于非天然前体构建目标产品的正交化人工合成途径[图3(c),正交化途径3].例如,以NPP(neryl diphosphate)替代其异构体GPP作为单萜合成的前体,通过NPP合酶的引入和单萜合酶的定向进化,可构建正交化的橙花醇、桧烯、柠檬烯等单萜类产品的合成途径[108]. ...
... [105].在碳源利用方面,常设计构建非天然途径,或利用葡萄糖、油脂等天然碳源[图3(c),正交化途径1],或转化半纤维素水解产物(木糖、阿拉伯糖、半乳糖)等替代碳源[图3(c),正交化途径2],合成关键前体化合物.例如,合成1,4-丁二醇[107]和琥珀酸[105]的正交化途径.在前体供给方面,常基于非天然前体构建目标产品的正交化人工合成途径[图3(c),正交化途径3].例如,以NPP(neryl diphosphate)替代其异构体GPP作为单萜合成的前体,通过NPP合酶的引入和单萜合酶的定向进化,可构建正交化的橙花醇、桧烯、柠檬烯等单萜类产品的合成途径[108]. ...
... [105]的正交化途径.在前体供给方面,常基于非天然前体构建目标产品的正交化人工合成途径[图3(c),正交化途径3].例如,以NPP(neryl diphosphate)替代其异构体GPP作为单萜合成的前体,通过NPP合酶的引入和单萜合酶的定向进化,可构建正交化的橙花醇、桧烯、柠檬烯等单萜类产品的合成途径[108]. ...
Beware of metaphors: chasses and orthogonality in synthetic biology
1
2011
... 在物质代谢水平,代谢途径的相对正交化是指底物同化途径或产物合成途径与底盘内源代谢网络最多只以唯一的结点相连,以形成独立的模块,便于后续调控其代谢流通量[105,106].此种途径的正交化过程可视作对底盘生长和产物合成的解耦[105].在碳源利用方面,常设计构建非天然途径,或利用葡萄糖、油脂等天然碳源[图3(c),正交化途径1],或转化半纤维素水解产物(木糖、阿拉伯糖、半乳糖)等替代碳源[图3(c),正交化途径2],合成关键前体化合物.例如,合成1,4-丁二醇[107]和琥珀酸[105]的正交化途径.在前体供给方面,常基于非天然前体构建目标产品的正交化人工合成途径[图3(c),正交化途径3].例如,以NPP(neryl diphosphate)替代其异构体GPP作为单萜合成的前体,通过NPP合酶的引入和单萜合酶的定向进化,可构建正交化的橙花醇、桧烯、柠檬烯等单萜类产品的合成途径[108]. ...
Autonomous production of 1,4-butanediol via a de novo biosynthesis pathway in engineered Escherichia coli
1
2015
... 在物质代谢水平,代谢途径的相对正交化是指底物同化途径或产物合成途径与底盘内源代谢网络最多只以唯一的结点相连,以形成独立的模块,便于后续调控其代谢流通量[105,106].此种途径的正交化过程可视作对底盘生长和产物合成的解耦[105].在碳源利用方面,常设计构建非天然途径,或利用葡萄糖、油脂等天然碳源[图3(c),正交化途径1],或转化半纤维素水解产物(木糖、阿拉伯糖、半乳糖)等替代碳源[图3(c),正交化途径2],合成关键前体化合物.例如,合成1,4-丁二醇[107]和琥珀酸[105]的正交化途径.在前体供给方面,常基于非天然前体构建目标产品的正交化人工合成途径[图3(c),正交化途径3].例如,以NPP(neryl diphosphate)替代其异构体GPP作为单萜合成的前体,通过NPP合酶的引入和单萜合酶的定向进化,可构建正交化的橙花醇、桧烯、柠檬烯等单萜类产品的合成途径[108]. ...
Orthogonal monoterpenoid biosynthesis in yeast constructed on an isomeric substrate
1
2019
... 在物质代谢水平,代谢途径的相对正交化是指底物同化途径或产物合成途径与底盘内源代谢网络最多只以唯一的结点相连,以形成独立的模块,便于后续调控其代谢流通量[105,106].此种途径的正交化过程可视作对底盘生长和产物合成的解耦[105].在碳源利用方面,常设计构建非天然途径,或利用葡萄糖、油脂等天然碳源[图3(c),正交化途径1],或转化半纤维素水解产物(木糖、阿拉伯糖、半乳糖)等替代碳源[图3(c),正交化途径2],合成关键前体化合物.例如,合成1,4-丁二醇[107]和琥珀酸[105]的正交化途径.在前体供给方面,常基于非天然前体构建目标产品的正交化人工合成途径[图3(c),正交化途径3].例如,以NPP(neryl diphosphate)替代其异构体GPP作为单萜合成的前体,通过NPP合酶的引入和单萜合酶的定向进化,可构建正交化的橙花醇、桧烯、柠檬烯等单萜类产品的合成途径[108]. ...
Creation of bioorthogonal redox systems depending on nicotinamide flucytosine dinucleotide
1
2011
... 物质转化和能量转移高度耦合.而天然辅酶同时参与众多代谢反应和生物过程,其具有选择性差、生物学效应可预见性低等问题[109],降低了所涉及途径的正交性.通过设计构建烟酰胺胞嘧啶二核苷酸等非天然辅酶[110,111,112],建立与底盘正交的氧化还原体系,可以从能量代谢的角度增强目标途径与内源代谢网络的正交化程度. ...
Structural insights into phosphite dehydrogenase variants favoring a non-natural redox cofactor
1
2019
... 物质转化和能量转移高度耦合.而天然辅酶同时参与众多代谢反应和生物过程,其具有选择性差、生物学效应可预见性低等问题[109],降低了所涉及途径的正交性.通过设计构建烟酰胺胞嘧啶二核苷酸等非天然辅酶[110,111,112],建立与底盘正交的氧化还原体系,可以从能量代谢的角度增强目标途径与内源代谢网络的正交化程度. ...
Synthetic cofactor-linked metabolic circuits for selective energy transfer
1
2017
... 物质转化和能量转移高度耦合.而天然辅酶同时参与众多代谢反应和生物过程,其具有选择性差、生物学效应可预见性低等问题[109],降低了所涉及途径的正交性.通过设计构建烟酰胺胞嘧啶二核苷酸等非天然辅酶[110,111,112],建立与底盘正交的氧化还原体系,可以从能量代谢的角度增强目标途径与内源代谢网络的正交化程度. ...
Non-natural cofactor and formate-driven reductive carboxylation of pyruvate
1
2020
... 物质转化和能量转移高度耦合.而天然辅酶同时参与众多代谢反应和生物过程,其具有选择性差、生物学效应可预见性低等问题[109],降低了所涉及途径的正交性.通过设计构建烟酰胺胞嘧啶二核苷酸等非天然辅酶[110,111,112],建立与底盘正交的氧化还原体系,可以从能量代谢的角度增强目标途径与内源代谢网络的正交化程度. ...
The yeast peroxisome: a dynamic storage depot and subcellular factory for squalene overproduction
1
2020
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Functional expression of a bacterial alpha-ketoglutarate dehydrogenase in the cytosol of Saccharomyces cerevisiae
1
2019
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Dual regulation of cytoplasmic and mitochondrial acetyl-CoA utilization for improved isoprene production in Saccharomyces cerevisiae
1
2016
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Cytosolic lipid droplets as engineered organelles for production and accumulation of terpenoid biomaterials in leaves
1
2019
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Co-compartmentation of terpene biosynthesis and storage via synthetic droplet
1
2018
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Harnessing yeast peroxisomes for biosynthesis of fatty-acid-derived biofuels and chemicals with relieved side-pathway competition
1
2016
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Tailoring the Saccharomyces cerevisiae endoplasmic reticulum for functional assembly of terpene synthesis pathway
1
2019
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Advanced strategies for production of natural products in yeast
1
2020
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Yeast peroxisomes: function and biogenesis of a versatile cell organelle
1
1997
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Towards designer organelles by subverting the peroxisomal import pathway
1
2017
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Controlled division of cell-sized vesicles by low densities of membrane-bound proteins
1
2020
... 与原核底盘相比,酿酒酵母等真核底盘具备多种可用于途径模块定位的细胞器,产生区室化效应[97],提高目标产物的合成水平[113,114,115,116,117].通过调控细胞器的形态、数量、大小等表型,可进一步提高定位途径的通量[118,119].如果靶向的细胞器具备正交性,可降低此种调控对其他细胞器及底盘内源代谢的干扰.根据参与中心代谢的程度,天然细胞器中过氧化物酶体的正交性最高[120],完全删除过氧化物酶体蛋白不会影响菌体以葡萄糖为底物的生长[121].而建立人工过氧化物酶体蛋白转运系统,在目标蛋白靶向定位的同时,抑制天然过氧化物酶体蛋白的定位表达[122],开启了定制化细胞器的研究[图3(d)].此外,借助低密度膜结合蛋白精确控制囊泡膜的自发弯曲,有望实现不依赖于底盘内源基因调控膜的分裂[123],而控制细胞器的表型. ...
Bacterial microcompartments
1
2018
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
Programmed loading and rapid purification of engineered bacterial microcompartment shells
1
2018
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
De novo targeting to the cytoplasmic and luminal side of bacterial microcompartments
1
2018
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
A designed bacterial microcompartment shell with tunable composition and precision cargo loading
1
2019
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
Widespread distribution of encapsulin nanocompartments reveals functional diversity
1
2017
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
Optogenetic protein clustering and signaling activation in mammalian cells
1
2013
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
Photoactivation of the BLUF protein PixD Probed by the site-specific incorporation of fluorotyrosine residues
1
2017
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
Prokaryotic nanocompartments form synthetic organelles in a eukaryote
1
2018
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
Light-based control of metabolic flux through assembly of synthetic organelles
1
2019
... 原核生物中天然存在由选择性通透的蛋白质衣壳代替磷脂层分子包裹形成的微室[124].在真核底盘中重建原核微室或构建类微室结构的人工细胞器,进行异源催化反应,也是细胞器正交化的策略手段[图3(d)].在重构原核微室时,采用SpyTag/SpyCatcher共价结合[125]或CC-Di-A/CC-Di-B卷曲螺旋(coiled-coil)[126]等蛋白质自主装结构,替代天然微室的包装序列,可解决催化蛋白易发生团聚而丧失活性的问题.但上述方法尚未在真核底盘中进行尝试[127].而参考天然微室结构,利用自组装蛋白(例如encapsulins[128]、受蓝光调控的Cry2[129]和PixD/PixE[130])的可控表达和组装,已成功在真核生物中构建了包装有催化蛋白的人工细胞器,实现norcoclaurine[131]、脱氧紫色杆菌素[132]等化合物的高效合成. ...
Overcoming genetic heterogeneity in industrial fermentations
1
2019
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Synthetic addiction extends the productive life time of engineered Escherichia coli populations
1
2018
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Exploiting nongenetic cell-to-cell variation for enhanced biosynthesis
1
2016
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Evolution-guided optimization of biosynthetic pathways
1
2014
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Dynamic regulation of metabolic flux in engineered bacteria using a pathway-independent quorum-sensing circuit
1
2017
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Engineering dynamic pathway regulation using stress-response promoters
2
2013
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
... [138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Dynamic control of toxic natural product biosynthesis by an artificial regulatory circuit
1
2020
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Burden-driven feedback control of gene expression
1
2018
... 构建基因线路,随菌体生长代谢或环境条件的变化对途径模块进行动态调控,是在代谢胁迫扰动下提高系统鲁棒性的有效手段[133].相关基因线路的设计原则一是偶联生长和生产[图3(e)],即产物在胞内的积累正反馈激活必需基因的表达,以避免负向突变在群体中的积累,强迫系统保持目标途径代谢流的稳定和最大化[134,135].在此基础上进一步采用同时具备正/负两向筛选功能的报告基因,可排除启动子区突变而产生的调控逃逸[136].另一基因线路设计原则是解耦生长和生产[图3(e)],基于群体响应元件,感知菌体的生长密度,反馈调控产物合成[137];或基于结合化学分子的调控蛋白,响应胞内关键代谢中间体的含量,实时调整产物的积累速率[35],防止有毒中间产物的过量积累[138],以降低异源途径对底盘的代谢胁迫.目前,调控蛋白从特异性识别一种化合物,发展为以不同的响应阈值同时识别两种以上化合物.在发酵前期通过底物的抑制作用限制有毒终产物的积累,在发酵后期通过产物的正反馈作用加快产物的合成[139].同时,以特异性响应某种代谢物的启动子可代替调控蛋白控制途径模块的表达[138].在此基础上级联dCas9/sgRNA系统的调控,还可增强响应速度和敏感性[140]. ...
Impact of synthetic biology and metabolic engineering on industrial production of fine chemicals
1
2015
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
The emergence of adaptive laboratory evolution as an efficient tool for biological discovery and industrial biotechnology
1
2019
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Evolutionary engineering reveals divergent paths when yeast is adapted to different acidic environments
1
2017
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Specificity of genome evolution in experimental populations of Escherichia coli evolved at different temperatures
1
2017
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Increased production of L-serine in Escherichia coli through adaptive laboratory evolution
1
2017
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Different tradeoffs result from alternate genetic adaptations to a common environment
1
2014
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Altered sterol composition renders yeast thermotolerant
1
2014
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Stress-tolerant non-conventional microbes enable next-generation chemical biosynthesis
1
2020
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Chasing bacterial chassis for metabolic engineering: a perspective review from classical to non-traditional microorganisms
1
2019
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Potential and limitations of Klebsiella pneumoniae as a microbial cell factory utilizing glycerol as the carbon source
1
2018
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Engineering Clostridium organisms as microbial cell-factories: challenges & opportunities
1
2018
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Engineering Halomonas TD01 for the low-cost production of polyhydroxyalkanoates
1
2014
... 为应对复杂、非理想化的工业发酵过程,需要底盘菌株拥有抗高温、耐盐、耐酸、耐有机溶剂等性状,以具备对某一类特定环境压力较高的鲁棒性[141].由于产生耐受性性状具有多基因性,且分子机制并未完全清晰阐释,现阶段很难通过工程化手段完整复制到大肠杆菌和酿酒酵母等常用模式底盘生物中.因此,多采用实验室适应性进化[142]等手段改善模式菌株的耐受性[143,144,145],并挖掘应对相关环境压力的基因组扰动靶点[146,147].近年来则倾向于选择天然对极端环境具有耐受性的非模式生物[148],逐步驯化为可用于细胞工厂构建的底盘菌株[149].例如耐热耐酸的马克斯克鲁维酵母[150]和溶剂耐受的丙酮丁醇梭菌[151].以嗜盐假单胞菌为底盘菌株,利用海水为介质高效生产聚羟基脂肪酸酯(PHA)等生物单体材料[152],是非模式底盘菌株工业应用的典型案例. ...
Engineering microbial consortia: a new frontier in synthetic biology
1
2008
... 有目标的设计、改造乃至重新合成微生物多细胞体系,在维持菌群结构与功能稳定性、鲁棒性和可控性的同时,可实现复杂的生物功能,提高代谢效率.人工多细胞体系在完成多项复杂任务方面优势明显,减轻单个底盘细胞的代谢负担,同时降低中间代谢物的过度积累和毒害,避免功能间的交叉影响,对环境波动具有更强的适应性和鲁棒性[153,154].因此,人工多细胞体系已成为合成生物学研究的重要方向[155,156]. ...
Use of bacterial co-cultures for the efficient production of chemicals
1
2018
... 有目标的设计、改造乃至重新合成微生物多细胞体系,在维持菌群结构与功能稳定性、鲁棒性和可控性的同时,可实现复杂的生物功能,提高代谢效率.人工多细胞体系在完成多项复杂任务方面优势明显,减轻单个底盘细胞的代谢负担,同时降低中间代谢物的过度积累和毒害,避免功能间的交叉影响,对环境波动具有更强的适应性和鲁棒性[153,154].因此,人工多细胞体系已成为合成生物学研究的重要方向[155,156]. ...
Synthetic microbial consortia: from systematic analysis to construction and applications
2
2014
... 有目标的设计、改造乃至重新合成微生物多细胞体系,在维持菌群结构与功能稳定性、鲁棒性和可控性的同时,可实现复杂的生物功能,提高代谢效率.人工多细胞体系在完成多项复杂任务方面优势明显,减轻单个底盘细胞的代谢负担,同时降低中间代谢物的过度积累和毒害,避免功能间的交叉影响,对环境波动具有更强的适应性和鲁棒性[153,154].因此,人工多细胞体系已成为合成生物学研究的重要方向[155,156]. ...
... 依据微生物间的互作关系设计构建人工多细胞体系,进一步借助代谢物互补、克服从底物到产物的能量壁垒、解除环境中的抑制作用、调控电子转移平衡等方式,使体系的物质流、能量流和信息流达到适配,可实现人工多细胞体系的高效稳定[155,157].细胞间互作关系的稳定存在,普遍依赖于细胞间通讯,细胞间通讯系统是构建人工多细胞体系的关键[158].研究者通过建立一种预测多细胞体系中相互作用关系的方法,可有效预测800多种菌群中互作关系[159].哈佛大学研究人员关注由细胞群体感应信号或代谢互补联系起来的混菌共生关系[160].利用群体感应系统,可构建能够双向通讯的多细胞体系[161]. ...
Better together: engineering and application of microbial symbioses
1
2015
... 有目标的设计、改造乃至重新合成微生物多细胞体系,在维持菌群结构与功能稳定性、鲁棒性和可控性的同时,可实现复杂的生物功能,提高代谢效率.人工多细胞体系在完成多项复杂任务方面优势明显,减轻单个底盘细胞的代谢负担,同时降低中间代谢物的过度积累和毒害,避免功能间的交叉影响,对环境波动具有更强的适应性和鲁棒性[153,154].因此,人工多细胞体系已成为合成生物学研究的重要方向[155,156]. ...
Designing microbial consortia with defined social interactions
1
2018
... 依据微生物间的互作关系设计构建人工多细胞体系,进一步借助代谢物互补、克服从底物到产物的能量壁垒、解除环境中的抑制作用、调控电子转移平衡等方式,使体系的物质流、能量流和信息流达到适配,可实现人工多细胞体系的高效稳定[155,157].细胞间互作关系的稳定存在,普遍依赖于细胞间通讯,细胞间通讯系统是构建人工多细胞体系的关键[158].研究者通过建立一种预测多细胞体系中相互作用关系的方法,可有效预测800多种菌群中互作关系[159].哈佛大学研究人员关注由细胞群体感应信号或代谢互补联系起来的混菌共生关系[160].利用群体感应系统,可构建能够双向通讯的多细胞体系[161]. ...
Cross-kingdom chemical communication drives a heritable, mutually beneficial prion-based transformation of metabolism
1
2014
... 依据微生物间的互作关系设计构建人工多细胞体系,进一步借助代谢物互补、克服从底物到产物的能量壁垒、解除环境中的抑制作用、调控电子转移平衡等方式,使体系的物质流、能量流和信息流达到适配,可实现人工多细胞体系的高效稳定[155,157].细胞间互作关系的稳定存在,普遍依赖于细胞间通讯,细胞间通讯系统是构建人工多细胞体系的关键[158].研究者通过建立一种预测多细胞体系中相互作用关系的方法,可有效预测800多种菌群中互作关系[159].哈佛大学研究人员关注由细胞群体感应信号或代谢互补联系起来的混菌共生关系[160].利用群体感应系统,可构建能够双向通讯的多细胞体系[161]. ...
Metabolic dependencies drive species co-occurrence in diverse microbial communities
1
2015
... 依据微生物间的互作关系设计构建人工多细胞体系,进一步借助代谢物互补、克服从底物到产物的能量壁垒、解除环境中的抑制作用、调控电子转移平衡等方式,使体系的物质流、能量流和信息流达到适配,可实现人工多细胞体系的高效稳定[155,157].细胞间互作关系的稳定存在,普遍依赖于细胞间通讯,细胞间通讯系统是构建人工多细胞体系的关键[158].研究者通过建立一种预测多细胞体系中相互作用关系的方法,可有效预测800多种菌群中互作关系[159].哈佛大学研究人员关注由细胞群体感应信号或代谢互补联系起来的混菌共生关系[160].利用群体感应系统,可构建能够双向通讯的多细胞体系[161]. ...
Dynamics in the mixed microbial concourse
1
2010
... 依据微生物间的互作关系设计构建人工多细胞体系,进一步借助代谢物互补、克服从底物到产物的能量壁垒、解除环境中的抑制作用、调控电子转移平衡等方式,使体系的物质流、能量流和信息流达到适配,可实现人工多细胞体系的高效稳定[155,157].细胞间互作关系的稳定存在,普遍依赖于细胞间通讯,细胞间通讯系统是构建人工多细胞体系的关键[158].研究者通过建立一种预测多细胞体系中相互作用关系的方法,可有效预测800多种菌群中互作关系[159].哈佛大学研究人员关注由细胞群体感应信号或代谢互补联系起来的混菌共生关系[160].利用群体感应系统,可构建能够双向通讯的多细胞体系[161]. ...
Engineered bidirectional communication mediates a consensus in a microbial biofilm consortium
1
2007
... 依据微生物间的互作关系设计构建人工多细胞体系,进一步借助代谢物互补、克服从底物到产物的能量壁垒、解除环境中的抑制作用、调控电子转移平衡等方式,使体系的物质流、能量流和信息流达到适配,可实现人工多细胞体系的高效稳定[155,157].细胞间互作关系的稳定存在,普遍依赖于细胞间通讯,细胞间通讯系统是构建人工多细胞体系的关键[158].研究者通过建立一种预测多细胞体系中相互作用关系的方法,可有效预测800多种菌群中互作关系[159].哈佛大学研究人员关注由细胞群体感应信号或代谢互补联系起来的混菌共生关系[160].利用群体感应系统,可构建能够双向通讯的多细胞体系[161]. ...
Microbial interactions: from networks to models
1
2012
... 自然条件下的多细胞体系,常形成互利、偏利、偏害、竞争等多种相互作用关系[162].自然形成或人工复配的多细胞体系中一般存在成员分工不明确、代谢水平不平衡、竞争与协作共存等问题,相互作用关系错综复杂.采用模块化构建的方法,可针对性地设计和重构各个功能菌种,构建菌株间的协作关系,使其有利于多细胞体系的高效性和稳定性[163,164]. ...
Synthetic biology tools to engineer microbial communities for biotechnology
2
2019
... 自然条件下的多细胞体系,常形成互利、偏利、偏害、竞争等多种相互作用关系[162].自然形成或人工复配的多细胞体系中一般存在成员分工不明确、代谢水平不平衡、竞争与协作共存等问题,相互作用关系错综复杂.采用模块化构建的方法,可针对性地设计和重构各个功能菌种,构建菌株间的协作关系,使其有利于多细胞体系的高效性和稳定性[163,164]. ...
... 近年来,人工合成多细胞体系的设计构建取得了重要研究进展,微生物混菌体系在资源能源、环境和人类健康领域等表现出强大的应用前景[163,177,178]. ...
A stabilized microbial ecosystem of self-limiting bacteria using synthetic quorum-regulated
1
2017
... 自然条件下的多细胞体系,常形成互利、偏利、偏害、竞争等多种相互作用关系[162].自然形成或人工复配的多细胞体系中一般存在成员分工不明确、代谢水平不平衡、竞争与协作共存等问题,相互作用关系错综复杂.采用模块化构建的方法,可针对性地设计和重构各个功能菌种,构建菌株间的协作关系,使其有利于多细胞体系的高效性和稳定性[163,164]. ...
Synthetic cooperation in engineered yeast populations
1
2007
... 在互利共生体系中,菌株之间共享或者交换营养物质,彼此之间可共同完成某一种化合物的彻底代谢,并分别从中获益.通过构建了营养缺陷型菌株,可以构建互利共生的多细胞系统,使菌株促进彼此的生长[165].研究者利用双向通讯在两株大肠杆菌中构建了共生系统[166].在偏利或偏害关系的体系中,对一方菌株有益/有害,另一方则不受影响.菌株间对环境资源(如营养物质、空间等)的争夺则形成了菌株间的竞争关系.在构建人工多细胞体系时,也应充分考虑菌株间竞争、偏利或偏害等关系存在的可能性.部分菌株可能加速对营养物质的吸收,并对这些物质进行与构建目标不相关的无效转化,或者产生对其他菌株有抑制作用的物质,这必然会影响菌群的高效和稳定.通过对混菌体系中菌株关系的重构,可解除竞争抑制,使菌株间偏利共生和竞争关系转变为互利共生[167].通过人工构建和调控菌株间的互利共生关系,使人工多细胞体系对复杂功能网络进行有效的分工协作,可实现混菌系统的高效、稳定、可控.此外,研究者还设计构建了多种菌群相互作用关系,为构建人工多细胞体系提供新的思路.例如:在两株大肠杆菌中分别构建群体感应信号调控下的细胞自杀基因表达及其抑制途径,可形成捕食者与被捕食者的关系[30]. ...
An environment-sensitive synthetic microbial ecosystem
1
2010
... 在互利共生体系中,菌株之间共享或者交换营养物质,彼此之间可共同完成某一种化合物的彻底代谢,并分别从中获益.通过构建了营养缺陷型菌株,可以构建互利共生的多细胞系统,使菌株促进彼此的生长[165].研究者利用双向通讯在两株大肠杆菌中构建了共生系统[166].在偏利或偏害关系的体系中,对一方菌株有益/有害,另一方则不受影响.菌株间对环境资源(如营养物质、空间等)的争夺则形成了菌株间的竞争关系.在构建人工多细胞体系时,也应充分考虑菌株间竞争、偏利或偏害等关系存在的可能性.部分菌株可能加速对营养物质的吸收,并对这些物质进行与构建目标不相关的无效转化,或者产生对其他菌株有抑制作用的物质,这必然会影响菌群的高效和稳定.通过对混菌体系中菌株关系的重构,可解除竞争抑制,使菌株间偏利共生和竞争关系转变为互利共生[167].通过人工构建和调控菌株间的互利共生关系,使人工多细胞体系对复杂功能网络进行有效的分工协作,可实现混菌系统的高效、稳定、可控.此外,研究者还设计构建了多种菌群相互作用关系,为构建人工多细胞体系提供新的思路.例如:在两株大肠杆菌中分别构建群体感应信号调控下的细胞自杀基因表达及其抑制途径,可形成捕食者与被捕食者的关系[30]. ...
Reorganization of a synthetic microbial consortium for one-step vitamin C fermentation
1
2016
... 在互利共生体系中,菌株之间共享或者交换营养物质,彼此之间可共同完成某一种化合物的彻底代谢,并分别从中获益.通过构建了营养缺陷型菌株,可以构建互利共生的多细胞系统,使菌株促进彼此的生长[165].研究者利用双向通讯在两株大肠杆菌中构建了共生系统[166].在偏利或偏害关系的体系中,对一方菌株有益/有害,另一方则不受影响.菌株间对环境资源(如营养物质、空间等)的争夺则形成了菌株间的竞争关系.在构建人工多细胞体系时,也应充分考虑菌株间竞争、偏利或偏害等关系存在的可能性.部分菌株可能加速对营养物质的吸收,并对这些物质进行与构建目标不相关的无效转化,或者产生对其他菌株有抑制作用的物质,这必然会影响菌群的高效和稳定.通过对混菌体系中菌株关系的重构,可解除竞争抑制,使菌株间偏利共生和竞争关系转变为互利共生[167].通过人工构建和调控菌株间的互利共生关系,使人工多细胞体系对复杂功能网络进行有效的分工协作,可实现混菌系统的高效、稳定、可控.此外,研究者还设计构建了多种菌群相互作用关系,为构建人工多细胞体系提供新的思路.例如:在两株大肠杆菌中分别构建群体感应信号调控下的细胞自杀基因表达及其抑制途径,可形成捕食者与被捕食者的关系[30]. ...
Engineering microbial consortia by division of labor
1
2019
... 依据劳动分工原则构建人工多细胞体系,可将复杂的任务合理分配给不同的菌株,在一定程度上避免单菌中细胞代谢负荷重等问题,在提高效率等方面具有显著优势[168].依据“劳动分工合作”的原则,构建出大肠杆菌-枯草芽孢杆菌-希瓦氏菌人工三菌产电体系,显著提高了化学能到电能的转化效率[169];通过将紫杉醇前体(5α-乙酰化-10β-羟化紫杉二烯)的合成途径分工构建到大肠杆菌-酿酒酵母人工两菌体系中,实现了目标产物的高效合成[170];为实现黄酮类化合物的合成,研究者设计构建大肠杆菌-大肠杆菌双菌体系,显著降低了单菌的代谢负担,使黄烷-3-醇的产量比单菌体系提高970倍[171]. ...
A three-species microbial consortium for power generation
2
2017
... 依据劳动分工原则构建人工多细胞体系,可将复杂的任务合理分配给不同的菌株,在一定程度上避免单菌中细胞代谢负荷重等问题,在提高效率等方面具有显著优势[168].依据“劳动分工合作”的原则,构建出大肠杆菌-枯草芽孢杆菌-希瓦氏菌人工三菌产电体系,显著提高了化学能到电能的转化效率[169];通过将紫杉醇前体(5α-乙酰化-10β-羟化紫杉二烯)的合成途径分工构建到大肠杆菌-酿酒酵母人工两菌体系中,实现了目标产物的高效合成[170];为实现黄酮类化合物的合成,研究者设计构建大肠杆菌-大肠杆菌双菌体系,显著降低了单菌的代谢负担,使黄烷-3-醇的产量比单菌体系提高970倍[171]. ...
... 近年来,人工三菌、四菌体系的构建和应用研究也取得重要进展.通过调控电子载体的合成、碳源的优化分配和利用,构建了高效稳定的人工三菌产电体系,能够利用0.28g葡萄糖稳定产电超过15天[169].通过构建和调控大肠杆菌三菌体系,减少副产物的形成,实现了木质素单体的高效合成[175].通过在四株大肠杆菌中表达15个外源基因,将天然产物花青素的合成途径分工构建到大肠杆菌四菌体系,首次实现了以葡萄糖、甘油和木糖为底物合成花青素[176]. ...
Distributing a metabolic pathway among a microbial consortium enhances production of natural products
1
2015
... 依据劳动分工原则构建人工多细胞体系,可将复杂的任务合理分配给不同的菌株,在一定程度上避免单菌中细胞代谢负荷重等问题,在提高效率等方面具有显著优势[168].依据“劳动分工合作”的原则,构建出大肠杆菌-枯草芽孢杆菌-希瓦氏菌人工三菌产电体系,显著提高了化学能到电能的转化效率[169];通过将紫杉醇前体(5α-乙酰化-10β-羟化紫杉二烯)的合成途径分工构建到大肠杆菌-酿酒酵母人工两菌体系中,实现了目标产物的高效合成[170];为实现黄酮类化合物的合成,研究者设计构建大肠杆菌-大肠杆菌双菌体系,显著降低了单菌的代谢负担,使黄烷-3-醇的产量比单菌体系提高970倍[171]. ...
Experimental and computational optimization of an Escherichia coli co-culture for the efficient production of flavonoids
2
2016
... 依据劳动分工原则构建人工多细胞体系,可将复杂的任务合理分配给不同的菌株,在一定程度上避免单菌中细胞代谢负荷重等问题,在提高效率等方面具有显著优势[168].依据“劳动分工合作”的原则,构建出大肠杆菌-枯草芽孢杆菌-希瓦氏菌人工三菌产电体系,显著提高了化学能到电能的转化效率[169];通过将紫杉醇前体(5α-乙酰化-10β-羟化紫杉二烯)的合成途径分工构建到大肠杆菌-酿酒酵母人工两菌体系中,实现了目标产物的高效合成[170];为实现黄酮类化合物的合成,研究者设计构建大肠杆菌-大肠杆菌双菌体系,显著降低了单菌的代谢负担,使黄烷-3-醇的产量比单菌体系提高970倍[171]. ...
... 在合成化学品、高附加值产品等方面,研究者通过构建大肠杆菌混菌体系,实现了黏康酸[179]、红景天苷[180]、丁醇[181]、吡喃花青素[182]、黄酮类化合物[171]、苯酚[183]等高效合成.如人工构建的大肠杆菌-大肠杆菌“营养互补”体系可以代谢葡萄糖和木糖生产红景天苷,通过两菌株的分工合成,红景天苷的混菌发酵单位达到6.03g/L,是单菌产量的20倍[180]. ...
Metabolic cross-feeding via intercellular nanotubes among bacteria
1
2015
... 人工多细胞体系中菌株间的互作交流主要有两种方式:一种是依赖于微生物细胞与细胞间的直接接触作用方式,主要包括膜囊泡、纳米管和瞬时融合外膜.如通过构建不动杆菌-大肠杆菌互利共生混菌体系研究两菌的代谢交流,发现两菌通过细菌纳米管进行物质交换和信息交流[172].另一种是菌株可通过间接接触进行物质和能量交换,以维持菌群的生态稳定性[173].间接接触主要依赖于释放到环境中的化学分子(蛋白质、代谢物等)和电子等.如可利用乙醛的挥发特性作为信号分子,构建不同菌株的共生关系[174]等. ...
Nutritional stress induces exchange of cell material and energetic coupling between bacterial species
1
2015
... 人工多细胞体系中菌株间的互作交流主要有两种方式:一种是依赖于微生物细胞与细胞间的直接接触作用方式,主要包括膜囊泡、纳米管和瞬时融合外膜.如通过构建不动杆菌-大肠杆菌互利共生混菌体系研究两菌的代谢交流,发现两菌通过细菌纳米管进行物质交换和信息交流[172].另一种是菌株可通过间接接触进行物质和能量交换,以维持菌群的生态稳定性[173].间接接触主要依赖于释放到环境中的化学分子(蛋白质、代谢物等)和电子等.如可利用乙醛的挥发特性作为信号分子,构建不同菌株的共生关系[174]等. ...
Synthetic ecosystems based on airborne inter- and intrakingdom communication
1
2007
... 人工多细胞体系中菌株间的互作交流主要有两种方式:一种是依赖于微生物细胞与细胞间的直接接触作用方式,主要包括膜囊泡、纳米管和瞬时融合外膜.如通过构建不动杆菌-大肠杆菌互利共生混菌体系研究两菌的代谢交流,发现两菌通过细菌纳米管进行物质交换和信息交流[172].另一种是菌株可通过间接接触进行物质和能量交换,以维持菌群的生态稳定性[173].间接接触主要依赖于释放到环境中的化学分子(蛋白质、代谢物等)和电子等.如可利用乙醛的挥发特性作为信号分子,构建不同菌株的共生关系[174]等. ...
Metabolic engineering of Escherichia coli for microbial synthesis of monolignols
1
2017
... 近年来,人工三菌、四菌体系的构建和应用研究也取得重要进展.通过调控电子载体的合成、碳源的优化分配和利用,构建了高效稳定的人工三菌产电体系,能够利用0.28g葡萄糖稳定产电超过15天[169].通过构建和调控大肠杆菌三菌体系,减少副产物的形成,实现了木质素单体的高效合成[175].通过在四株大肠杆菌中表达15个外源基因,将天然产物花青素的合成途径分工构建到大肠杆菌四菌体系,首次实现了以葡萄糖、甘油和木糖为底物合成花青素[176]. ...
Complete biosynthesis of Anthocyanins using E. coli polycultures
1
2017
... 近年来,人工三菌、四菌体系的构建和应用研究也取得重要进展.通过调控电子载体的合成、碳源的优化分配和利用,构建了高效稳定的人工三菌产电体系,能够利用0.28g葡萄糖稳定产电超过15天[169].通过构建和调控大肠杆菌三菌体系,减少副产物的形成,实现了木质素单体的高效合成[175].通过在四株大肠杆菌中表达15个外源基因,将天然产物花青素的合成途径分工构建到大肠杆菌四菌体系,首次实现了以葡萄糖、甘油和木糖为底物合成花青素[176]. ...
Biotechnological potential and applications of microbial consortia
1
2020
... 近年来,人工合成多细胞体系的设计构建取得了重要研究进展,微生物混菌体系在资源能源、环境和人类健康领域等表现出强大的应用前景[163,177,178]. ...
Synthetic microbial consortia for small molecule production
1
2019
... 近年来,人工合成多细胞体系的设计构建取得了重要研究进展,微生物混菌体系在资源能源、环境和人类健康领域等表现出强大的应用前景[163,177,178]. ...
Engineering Escherichia coli coculture systems for the production of biochemical products
1
2015
... 在合成化学品、高附加值产品等方面,研究者通过构建大肠杆菌混菌体系,实现了黏康酸[179]、红景天苷[180]、丁醇[181]、吡喃花青素[182]、黄酮类化合物[171]、苯酚[183]等高效合成.如人工构建的大肠杆菌-大肠杆菌“营养互补”体系可以代谢葡萄糖和木糖生产红景天苷,通过两菌株的分工合成,红景天苷的混菌发酵单位达到6.03g/L,是单菌产量的20倍[180]. ...
Convergent engineering of syntrophic Escherichia coli coculture for efficient production of glycosides
2
2018
... 在合成化学品、高附加值产品等方面,研究者通过构建大肠杆菌混菌体系,实现了黏康酸[179]、红景天苷[180]、丁醇[181]、吡喃花青素[182]、黄酮类化合物[171]、苯酚[183]等高效合成.如人工构建的大肠杆菌-大肠杆菌“营养互补”体系可以代谢葡萄糖和木糖生产红景天苷,通过两菌株的分工合成,红景天苷的混菌发酵单位达到6.03g/L,是单菌产量的20倍[180]. ...
... [180]. ...
Design and development of a "Y-shaped" microbial consortium capable of simultaneously utilizing biomass sugars for efficient production of butanol
1
2019
... 在合成化学品、高附加值产品等方面,研究者通过构建大肠杆菌混菌体系,实现了黏康酸[179]、红景天苷[180]、丁醇[181]、吡喃花青素[182]、黄酮类化合物[171]、苯酚[183]等高效合成.如人工构建的大肠杆菌-大肠杆菌“营养互补”体系可以代谢葡萄糖和木糖生产红景天苷,通过两菌株的分工合成,红景天苷的混菌发酵单位达到6.03g/L,是单菌产量的20倍[180]. ...
Production of pyranoanthocyanins using Escherichia coli co-cultures
1
2019
... 在合成化学品、高附加值产品等方面,研究者通过构建大肠杆菌混菌体系,实现了黏康酸[179]、红景天苷[180]、丁醇[181]、吡喃花青素[182]、黄酮类化合物[171]、苯酚[183]等高效合成.如人工构建的大肠杆菌-大肠杆菌“营养互补”体系可以代谢葡萄糖和木糖生产红景天苷,通过两菌株的分工合成,红景天苷的混菌发酵单位达到6.03g/L,是单菌产量的20倍[180]. ...
De novo phenol bioproduction from glucose using biosensor-assisted microbial coculture engineering
1
2019
... 在合成化学品、高附加值产品等方面,研究者通过构建大肠杆菌混菌体系,实现了黏康酸[179]、红景天苷[180]、丁醇[181]、吡喃花青素[182]、黄酮类化合物[171]、苯酚[183]等高效合成.如人工构建的大肠杆菌-大肠杆菌“营养互补”体系可以代谢葡萄糖和木糖生产红景天苷,通过两菌株的分工合成,红景天苷的混菌发酵单位达到6.03g/L,是单菌产量的20倍[180]. ...
Design and characterization of synthetic fungal-bacterial consortia for direct production of isobutanol from cellulosic biomass
1
2013
... 在纤维素等复杂化合物的降解方面,通过调控两菌体系的菌群关系和菌群结构,实现了高效转化木质纤维素生产丁醇[184]和富马酸[185];通过在酵母表面展示纤维小体形成四菌体系,有效利用纤维素生产乙醇[186];通过增强“营养-解毒”关系,构建了双梭菌互利共生体系,可将玉米芯转化为丁醇,溶剂终浓度达到22g/L[187]. ...
Production of cellulosic organic acids via synthetic fungal consortia
1
2018
... 在纤维素等复杂化合物的降解方面,通过调控两菌体系的菌群关系和菌群结构,实现了高效转化木质纤维素生产丁醇[184]和富马酸[185];通过在酵母表面展示纤维小体形成四菌体系,有效利用纤维素生产乙醇[186];通过增强“营养-解毒”关系,构建了双梭菌互利共生体系,可将玉米芯转化为丁醇,溶剂终浓度达到22g/L[187]. ...
Surface display of a functional minicellulosome by intracellular complementation using a synthetic yeast consortium and its application to cellulose hydrolysis and ethanol production
1
2010
... 在纤维素等复杂化合物的降解方面,通过调控两菌体系的菌群关系和菌群结构,实现了高效转化木质纤维素生产丁醇[184]和富马酸[185];通过在酵母表面展示纤维小体形成四菌体系,有效利用纤维素生产乙醇[186];通过增强“营养-解毒”关系,构建了双梭菌互利共生体系,可将玉米芯转化为丁醇,溶剂终浓度达到22g/L[187]. ...
Enhanced solvent production by metabolic engineering of a twin-clostridial consortium
1
2017
... 在纤维素等复杂化合物的降解方面,通过调控两菌体系的菌群关系和菌群结构,实现了高效转化木质纤维素生产丁醇[184]和富马酸[185];通过在酵母表面展示纤维小体形成四菌体系,有效利用纤维素生产乙醇[186];通过增强“营养-解毒”关系,构建了双梭菌互利共生体系,可将玉米芯转化为丁醇,溶剂终浓度达到22g/L[187]. ...
A constructed microbial consortium for biodegradation of the organophosphorus insecticide parathion
1
2003
... 人工多细胞体系在环境污染物降解、土壤修复等方面也表现出巨大潜力.人工构建的由两株工程化大肠杆菌构成的混菌体系,实现了降解有机磷杀虫剂[188].人工构建的大肠杆菌-苍白杆菌混菌体系,可实现对二甲基有机磷的完全矿化[189].通过构建两株假单胞菌人工混菌体系,可提高对石油硫化物的脱硫作用[190].人工构建的四菌体系可用于降解碳氢化合物[191],如常见的原料(橄榄油、石蜡油等)和多芳烃污染物(萘和蒽)等,同时生产表面活性剂. ...
Functional assembly of a microbial consortium with autofluorescent and mineralizing activity for the biodegradation of organophosphates
1
2008
... 人工多细胞体系在环境污染物降解、土壤修复等方面也表现出巨大潜力.人工构建的由两株工程化大肠杆菌构成的混菌体系,实现了降解有机磷杀虫剂[188].人工构建的大肠杆菌-苍白杆菌混菌体系,可实现对二甲基有机磷的完全矿化[189].通过构建两株假单胞菌人工混菌体系,可提高对石油硫化物的脱硫作用[190].人工构建的四菌体系可用于降解碳氢化合物[191],如常见的原料(橄榄油、石蜡油等)和多芳烃污染物(萘和蒽)等,同时生产表面活性剂. ...
Engineering synthetic bacterial consortia for enhanced desulfurization and revalorization of oil sulfur compounds
1
2016
... 人工多细胞体系在环境污染物降解、土壤修复等方面也表现出巨大潜力.人工构建的由两株工程化大肠杆菌构成的混菌体系,实现了降解有机磷杀虫剂[188].人工构建的大肠杆菌-苍白杆菌混菌体系,可实现对二甲基有机磷的完全矿化[189].通过构建两株假单胞菌人工混菌体系,可提高对石油硫化物的脱硫作用[190].人工构建的四菌体系可用于降解碳氢化合物[191],如常见的原料(橄榄油、石蜡油等)和多芳烃污染物(萘和蒽)等,同时生产表面活性剂. ...
Construction of a hydrocarbon-degrading consortium and characterization of two new lipopeptides biosurfactants
1
2020
... 人工多细胞体系在环境污染物降解、土壤修复等方面也表现出巨大潜力.人工构建的由两株工程化大肠杆菌构成的混菌体系,实现了降解有机磷杀虫剂[188].人工构建的大肠杆菌-苍白杆菌混菌体系,可实现对二甲基有机磷的完全矿化[189].通过构建两株假单胞菌人工混菌体系,可提高对石油硫化物的脱硫作用[190].人工构建的四菌体系可用于降解碳氢化合物[191],如常见的原料(橄榄油、石蜡油等)和多芳烃污染物(萘和蒽)等,同时生产表面活性剂. ...
DNA Fountain enables a robust and efficient storage architecture
2
2017
... 手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中.随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战.受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略.DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据[192,193,194,195,196].DNA存储的核心优势是:①载体特征尺度小,信息存储密度高.例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB = 1015字节)的存储密度[192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
... [192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
Random access in large-scale DNA data storage
1
2018
... 手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中.随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战.受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略.DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据[192,193,194,195,196].DNA存储的核心优势是:①载体特征尺度小,信息存储密度高.例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB = 1015字节)的存储密度[192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
Molecular digital data storage using DNA
2
2019
... 手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中.随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战.受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略.DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据[192,193,194,195,196].DNA存储的核心优势是:①载体特征尺度小,信息存储密度高.例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB = 1015字节)的存储密度[192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
... [194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
Towards practical, high-capacity, low-maintenance information storage in synthesized DNA
1
2013
... 手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中.随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战.受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略.DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据[192,193,194,195,196].DNA存储的核心优势是:①载体特征尺度小,信息存储密度高.例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB = 1015字节)的存储密度[192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
Next-generation digital information storage in DNA
1
2012
... 手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中.随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战.受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略.DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据[192,193,194,195,196].DNA存储的核心优势是:①载体特征尺度小,信息存储密度高.例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB = 1015字节)的存储密度[192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
A DNA-of-things storage architecture to create materials with embedded memory
1
2020
... 手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中.随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战.受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略.DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据[192,193,194,195,196].DNA存储的核心优势是:①载体特征尺度小,信息存储密度高.例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB = 1015字节)的存储密度[192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
Robust chemical preservation of digital information on DNA in silica with error-correcting codes
1
2015
... 手指、石头、刻痕、绳结在古代被用来记录数据和信息,现在大量的信息主要存储于基于电、光和磁的存储介质中.随着社会发展,不断增长的数据存储需求面临着密度、功耗、体积等诸多挑战.受DNA存储遗传信息启发,人造DNA序列被认为是信息存储的新策略.DNA信息存储是先将数据进行编码转换为DNA序列,借助DNA合成数据存入DNA分子;信息读取是通过高通量测序和信息处理将DNA的分子序列转换为原始存储的数据[192,193,194,195,196].DNA存储的核心优势是:①载体特征尺度小,信息存储密度高.例如2017年,研究者利用数字信息传输中的喷泉编码实现了平均千分子DNA拷贝的数据存储与读取,实现了218 PB/g(1PB = 1015字节)的存储密度[192].微软等进一步优化信息存储DNA读取中PCR参数,实现了存储密度的进一步提升[194].②DNA可常温保存,维护成本显著降低,且保存时间久.瑞士苏黎世理工的研究者将DNA封存在特殊的材料中,有望存储千年甚至万年[197,198]. ...
Demonstration of end-to-end automation of DNA data storage
1
2019
... DNA存储作为一种新型数据存储方式,也面临很多发展挑战.一是传统的DNA合成技术主要针对生物学研究与应用建立,合成速度较低,成本较高,合成序列的长度与通量等也与数据存储需求存在很大差距.由于目前序列合成的成本远高于测序成本,DNA存储特别适合大量冷数据的长期存储.二是存储在DNA中的数据的读取灵活性仍有待提升,基于目前测序技术的读出方法面临数据恢复可靠性、仪器形态、环境适应性等诸多挑战.2019年微软开发了从写到读的端到端原型系统[199],距离现代信息存储系统仍有巨大发展空间.三是DNA信息存储与现有电子信息存储之间的编码转化尚不成熟.DNA作为一种物质介质具有各种自身的特点,如有断裂、突变、缺失等风险,近年来各种编码方法在DNA存储的密度、可靠性等方面有所突破[200,201,202].最后,DNA存储的技术链条较长,亟需生物、信息、仪器、机械和工程等领域的整合. ...
High information capacity DNA-based data storage with augmented encoding characters using degenerate bases
1
2019
... DNA存储作为一种新型数据存储方式,也面临很多发展挑战.一是传统的DNA合成技术主要针对生物学研究与应用建立,合成速度较低,成本较高,合成序列的长度与通量等也与数据存储需求存在很大差距.由于目前序列合成的成本远高于测序成本,DNA存储特别适合大量冷数据的长期存储.二是存储在DNA中的数据的读取灵活性仍有待提升,基于目前测序技术的读出方法面临数据恢复可靠性、仪器形态、环境适应性等诸多挑战.2019年微软开发了从写到读的端到端原型系统[199],距离现代信息存储系统仍有巨大发展空间.三是DNA信息存储与现有电子信息存储之间的编码转化尚不成熟.DNA作为一种物质介质具有各种自身的特点,如有断裂、突变、缺失等风险,近年来各种编码方法在DNA存储的密度、可靠性等方面有所突破[200,201,202].最后,DNA存储的技术链条较长,亟需生物、信息、仪器、机械和工程等领域的整合. ...
Data storage in DNA with fewer synthesis cycles using composite DNA letters
1
2019
... DNA存储作为一种新型数据存储方式,也面临很多发展挑战.一是传统的DNA合成技术主要针对生物学研究与应用建立,合成速度较低,成本较高,合成序列的长度与通量等也与数据存储需求存在很大差距.由于目前序列合成的成本远高于测序成本,DNA存储特别适合大量冷数据的长期存储.二是存储在DNA中的数据的读取灵活性仍有待提升,基于目前测序技术的读出方法面临数据恢复可靠性、仪器形态、环境适应性等诸多挑战.2019年微软开发了从写到读的端到端原型系统[199],距离现代信息存储系统仍有巨大发展空间.三是DNA信息存储与现有电子信息存储之间的编码转化尚不成熟.DNA作为一种物质介质具有各种自身的特点,如有断裂、突变、缺失等风险,近年来各种编码方法在DNA存储的密度、可靠性等方面有所突破[200,201,202].最后,DNA存储的技术链条较长,亟需生物、信息、仪器、机械和工程等领域的整合. ...
音视频文件的DNA信息存储
1
2020
... DNA存储作为一种新型数据存储方式,也面临很多发展挑战.一是传统的DNA合成技术主要针对生物学研究与应用建立,合成速度较低,成本较高,合成序列的长度与通量等也与数据存储需求存在很大差距.由于目前序列合成的成本远高于测序成本,DNA存储特别适合大量冷数据的长期存储.二是存储在DNA中的数据的读取灵活性仍有待提升,基于目前测序技术的读出方法面临数据恢复可靠性、仪器形态、环境适应性等诸多挑战.2019年微软开发了从写到读的端到端原型系统[199],距离现代信息存储系统仍有巨大发展空间.三是DNA信息存储与现有电子信息存储之间的编码转化尚不成熟.DNA作为一种物质介质具有各种自身的特点,如有断裂、突变、缺失等风险,近年来各种编码方法在DNA存储的密度、可靠性等方面有所突破[200,201,202].最后,DNA存储的技术链条较长,亟需生物、信息、仪器、机械和工程等领域的整合. ...
DNA information storage for audio and video files (in Chinese)
1
2020
... DNA存储作为一种新型数据存储方式,也面临很多发展挑战.一是传统的DNA合成技术主要针对生物学研究与应用建立,合成速度较低,成本较高,合成序列的长度与通量等也与数据存储需求存在很大差距.由于目前序列合成的成本远高于测序成本,DNA存储特别适合大量冷数据的长期存储.二是存储在DNA中的数据的读取灵活性仍有待提升,基于目前测序技术的读出方法面临数据恢复可靠性、仪器形态、环境适应性等诸多挑战.2019年微软开发了从写到读的端到端原型系统[199],距离现代信息存储系统仍有巨大发展空间.三是DNA信息存储与现有电子信息存储之间的编码转化尚不成熟.DNA作为一种物质介质具有各种自身的特点,如有断裂、突变、缺失等风险,近年来各种编码方法在DNA存储的密度、可靠性等方面有所突破[200,201,202].最后,DNA存储的技术链条较长,亟需生物、信息、仪器、机械和工程等领域的整合. ...
Folding DNA to create nanoscale shapes and patterns
1
2006
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
DNA纳米结构仿中国地图
1
2006
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
Imating China map with DNA nanostructure
1
2006
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
DNA-nanotube-induced alignment of membrane proteins for NMR structure determination
1
2007
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
Self-assembly of a nanoscale DNA box with a controllable lid
1
2009
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
DNA origami with complex curvatures in three-dimensional space
1
2011
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
DNA-nanostructure-gold-nanorod hybrids for enhanced in vivo optoacoustic imaging and photothermal therapy
1
2016
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
DNA origami cryptography for secure communication
1
2019
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
Tunable DNA origami motors translocate ballistically over μm distances at nm/s speeds
1
2020
... DNA作为纳米分子还可以被当作材料的构筑单元.DNA本身的特异性碱基互补配对为特异性组装材料提供了基础,可以构筑出以DNA作为框架的精细纳米结构.DNA折纸就是利用DNA的折叠和自组装形成复杂结构的技术,人工设计一系列DNA片段,借助碱基互补配对原则,可以折叠形成不同的预期几何结构,如三角形、矩形、菱形、五角星及笑脸等精细二维结构[203].我国学者利用人工设计DNA构建了微型的中国地图[204].其后,研究者还采用DNA折纸技术制造了齿轮、纳米盒子和花瓶等精细三维结构[205,206,207].由于采用了“自下而上”的理性组装方式,DNA折纸技术在纳米材料、分析科学、医药学、纳米机器人和信息加密等方面均有不同的创新性应用.特殊的DNA折纸结构载体可以用于包裹纳米金造影剂,显著提高成像质量,实现对肿瘤内部结构成像,极大减少造影剂的用量[208].DNA折纸技术还可以用于分子加密系统,性能可以超越基于硅基计算机的常规加密体系,且同时具有保护信息完整性和访问控制的功能[209].研究人员利用DNA折纸技术的精细可控性能,创建了迄今为止最快、最持久的DNA纳米马达[210]. ...
Beyond the canonical 20 amino acids: expanding the genetic lexicon
1
2010
... 天然蛋白质由20种天然氨基酸组成,迄今已有超过150种非天然氨基酸可以引入生物体内,其中大多数非天然氨基酸是天然氨基酸的衍生物.非天然氨基酸中根据目的需求可以引入烯基、酮基、醛基、酰氨基等功能基团,调控蛋白质的结构与功能[211].非天然氨基酸通过对特定蛋白质的修饰,可在生物催化、人工生物防逃逸、蛋白质药物、生物检测等方面有重要的应用.氟色氨酸的引入不仅可以提升转氨酶的活性,还提升了酶的溶剂耐受性和热稳定性[212].对氨基苯丙氨酸引入到转录因子蛋白LmrR可以将其改变为具有催化活性的腙和肟合成酶[213].此外,将非天然氨基酸引入大肠杆菌的必需蛋白,可以使大肠杆菌依赖于非天然氨基酸存活,从而达到人工生物体的生物安全防逃逸控制[214,215].卤代酪氨酸引入肌红蛋白氧化酶,可以形成光谱探针,为研究酶的催化机制提供了便捷手段[216]. ...
Enhancing thermostability and organic solvent tolerance of w-transaminase through global incorporation of fluorotyrosine
1
2014
... 天然蛋白质由20种天然氨基酸组成,迄今已有超过150种非天然氨基酸可以引入生物体内,其中大多数非天然氨基酸是天然氨基酸的衍生物.非天然氨基酸中根据目的需求可以引入烯基、酮基、醛基、酰氨基等功能基团,调控蛋白质的结构与功能[211].非天然氨基酸通过对特定蛋白质的修饰,可在生物催化、人工生物防逃逸、蛋白质药物、生物检测等方面有重要的应用.氟色氨酸的引入不仅可以提升转氨酶的活性,还提升了酶的溶剂耐受性和热稳定性[212].对氨基苯丙氨酸引入到转录因子蛋白LmrR可以将其改变为具有催化活性的腙和肟合成酶[213].此外,将非天然氨基酸引入大肠杆菌的必需蛋白,可以使大肠杆菌依赖于非天然氨基酸存活,从而达到人工生物体的生物安全防逃逸控制[214,215].卤代酪氨酸引入肌红蛋白氧化酶,可以形成光谱探针,为研究酶的催化机制提供了便捷手段[216]. ...
A designer enzyme for hydrazone and oxime formation featuring an unnatural catalytic aniline residue
1
2018
... 天然蛋白质由20种天然氨基酸组成,迄今已有超过150种非天然氨基酸可以引入生物体内,其中大多数非天然氨基酸是天然氨基酸的衍生物.非天然氨基酸中根据目的需求可以引入烯基、酮基、醛基、酰氨基等功能基团,调控蛋白质的结构与功能[211].非天然氨基酸通过对特定蛋白质的修饰,可在生物催化、人工生物防逃逸、蛋白质药物、生物检测等方面有重要的应用.氟色氨酸的引入不仅可以提升转氨酶的活性,还提升了酶的溶剂耐受性和热稳定性[212].对氨基苯丙氨酸引入到转录因子蛋白LmrR可以将其改变为具有催化活性的腙和肟合成酶[213].此外,将非天然氨基酸引入大肠杆菌的必需蛋白,可以使大肠杆菌依赖于非天然氨基酸存活,从而达到人工生物体的生物安全防逃逸控制[214,215].卤代酪氨酸引入肌红蛋白氧化酶,可以形成光谱探针,为研究酶的催化机制提供了便捷手段[216]. ...
Recoded organisms engineered to depend on synthetic amino acids
1
2015
... 天然蛋白质由20种天然氨基酸组成,迄今已有超过150种非天然氨基酸可以引入生物体内,其中大多数非天然氨基酸是天然氨基酸的衍生物.非天然氨基酸中根据目的需求可以引入烯基、酮基、醛基、酰氨基等功能基团,调控蛋白质的结构与功能[211].非天然氨基酸通过对特定蛋白质的修饰,可在生物催化、人工生物防逃逸、蛋白质药物、生物检测等方面有重要的应用.氟色氨酸的引入不仅可以提升转氨酶的活性,还提升了酶的溶剂耐受性和热稳定性[212].对氨基苯丙氨酸引入到转录因子蛋白LmrR可以将其改变为具有催化活性的腙和肟合成酶[213].此外,将非天然氨基酸引入大肠杆菌的必需蛋白,可以使大肠杆菌依赖于非天然氨基酸存活,从而达到人工生物体的生物安全防逃逸控制[214,215].卤代酪氨酸引入肌红蛋白氧化酶,可以形成光谱探针,为研究酶的催化机制提供了便捷手段[216]. ...
Biocontainment of genetically modified organisms by synthetic protein design
1
2015
... 天然蛋白质由20种天然氨基酸组成,迄今已有超过150种非天然氨基酸可以引入生物体内,其中大多数非天然氨基酸是天然氨基酸的衍生物.非天然氨基酸中根据目的需求可以引入烯基、酮基、醛基、酰氨基等功能基团,调控蛋白质的结构与功能[211].非天然氨基酸通过对特定蛋白质的修饰,可在生物催化、人工生物防逃逸、蛋白质药物、生物检测等方面有重要的应用.氟色氨酸的引入不仅可以提升转氨酶的活性,还提升了酶的溶剂耐受性和热稳定性[212].对氨基苯丙氨酸引入到转录因子蛋白LmrR可以将其改变为具有催化活性的腙和肟合成酶[213].此外,将非天然氨基酸引入大肠杆菌的必需蛋白,可以使大肠杆菌依赖于非天然氨基酸存活,从而达到人工生物体的生物安全防逃逸控制[214,215].卤代酪氨酸引入肌红蛋白氧化酶,可以形成光谱探针,为研究酶的催化机制提供了便捷手段[216]. ...
Defining the role of tyrosine and rational tuning of oxidase activity by genetic incorporation of unnatural tyrosine analogs
1
2015
... 天然蛋白质由20种天然氨基酸组成,迄今已有超过150种非天然氨基酸可以引入生物体内,其中大多数非天然氨基酸是天然氨基酸的衍生物.非天然氨基酸中根据目的需求可以引入烯基、酮基、醛基、酰氨基等功能基团,调控蛋白质的结构与功能[211].非天然氨基酸通过对特定蛋白质的修饰,可在生物催化、人工生物防逃逸、蛋白质药物、生物检测等方面有重要的应用.氟色氨酸的引入不仅可以提升转氨酶的活性,还提升了酶的溶剂耐受性和热稳定性[212].对氨基苯丙氨酸引入到转录因子蛋白LmrR可以将其改变为具有催化活性的腙和肟合成酶[213].此外,将非天然氨基酸引入大肠杆菌的必需蛋白,可以使大肠杆菌依赖于非天然氨基酸存活,从而达到人工生物体的生物安全防逃逸控制[214,215].卤代酪氨酸引入肌红蛋白氧化酶,可以形成光谱探针,为研究酶的催化机制提供了便捷手段[216]. ...
Playing with the molecules of life
1
2018
... 遗传密码拓展技术是非天然氨基酸引入的重要手段,主要是通过控制终止密码子的读取,进而通过氨酰-tRNA合成酶/tRNA对的改造实现[217].近年来基因组密码子删减研究可以为遗传密码拓展提供高效的人工底盘细胞[51,60],将明显提升引入非天然氨基酸的数量、种类和效率.此外,非天然核酸加入基因组的研究也将为更多非天然氨基酸的进一步高效加入提供新的方向[218]. ...
A semi-synthetic organism that stores and retrieves increased genetic information
3
2017
... 遗传密码拓展技术是非天然氨基酸引入的重要手段,主要是通过控制终止密码子的读取,进而通过氨酰-tRNA合成酶/tRNA对的改造实现[217].近年来基因组密码子删减研究可以为遗传密码拓展提供高效的人工底盘细胞[51,60],将明显提升引入非天然氨基酸的数量、种类和效率.此外,非天然核酸加入基因组的研究也将为更多非天然氨基酸的进一步高效加入提供新的方向[218]. ...
... 针对核酸的碱基、磷酸和五碳糖三个主要组成部分,可以分别进行人工修饰或替换,形成非天然核酸(XNA)这样一类核酸分子类似物[219].XNA能够通过碱基修饰或骨架修饰来拓展遗传信息的存储能力,可以具有部分或全部天然核酸的属性[220].生物体系引入XNA能够拓展遗传信息的多样性,进而拓展生物性能.非天然碱基研究近年来取得了一些重大突破,如八碱基DNA和具有生命活性的六碱基DNA等.通过对天然碱基(A、T、C、G)的分子结构进行修饰,创造出了两对新的可以互补配对的碱基(S、B、P、Z,其中S和B配对、P和Z配对).将人工碱基与天然碱基相结合,可以获得含有八种碱基的DNA[221].研究者在探索非天然碱基多样性的同时,也在尝试使非天然碱基具有生物活性.美国学者在大肠杆菌里成功表达了含有两个非天然碱基编码非天然氨基酸的荧光蛋白,实现了非天然碱基的转录活性,首次实现含六碱基DNA的半人工合成生命[218].XNA种类的增加不仅提升了遗传信息的密度,还为拓展生物的多样性奠定了基础. ...
... 目前XNA主要通过外源添加方式进入生物体[218],将来开发生物体内合成XNA,将进一步拓展XNA在合成生物学领域的广泛应用.由于XNA在体内的稳定性和特异性有其自身特点,在合成生物学的精准检测和正交控制等方面具有重要的应用潜力[222]. ...
Analysis of aptamer discovery and technology
1
2017
... 针对核酸的碱基、磷酸和五碳糖三个主要组成部分,可以分别进行人工修饰或替换,形成非天然核酸(XNA)这样一类核酸分子类似物[219].XNA能够通过碱基修饰或骨架修饰来拓展遗传信息的存储能力,可以具有部分或全部天然核酸的属性[220].生物体系引入XNA能够拓展遗传信息的多样性,进而拓展生物性能.非天然碱基研究近年来取得了一些重大突破,如八碱基DNA和具有生命活性的六碱基DNA等.通过对天然碱基(A、T、C、G)的分子结构进行修饰,创造出了两对新的可以互补配对的碱基(S、B、P、Z,其中S和B配对、P和Z配对).将人工碱基与天然碱基相结合,可以获得含有八种碱基的DNA[221].研究者在探索非天然碱基多样性的同时,也在尝试使非天然碱基具有生物活性.美国学者在大肠杆菌里成功表达了含有两个非天然碱基编码非天然氨基酸的荧光蛋白,实现了非天然碱基的转录活性,首次实现含六碱基DNA的半人工合成生命[218].XNA种类的增加不仅提升了遗传信息的密度,还为拓展生物的多样性奠定了基础. ...
The expanding world of DNA and RNA
1
2016
... 针对核酸的碱基、磷酸和五碳糖三个主要组成部分,可以分别进行人工修饰或替换,形成非天然核酸(XNA)这样一类核酸分子类似物[219].XNA能够通过碱基修饰或骨架修饰来拓展遗传信息的存储能力,可以具有部分或全部天然核酸的属性[220].生物体系引入XNA能够拓展遗传信息的多样性,进而拓展生物性能.非天然碱基研究近年来取得了一些重大突破,如八碱基DNA和具有生命活性的六碱基DNA等.通过对天然碱基(A、T、C、G)的分子结构进行修饰,创造出了两对新的可以互补配对的碱基(S、B、P、Z,其中S和B配对、P和Z配对).将人工碱基与天然碱基相结合,可以获得含有八种碱基的DNA[221].研究者在探索非天然碱基多样性的同时,也在尝试使非天然碱基具有生物活性.美国学者在大肠杆菌里成功表达了含有两个非天然碱基编码非天然氨基酸的荧光蛋白,实现了非天然碱基的转录活性,首次实现含六碱基DNA的半人工合成生命[218].XNA种类的增加不仅提升了遗传信息的密度,还为拓展生物的多样性奠定了基础. ...
Hachimoji DNA and RNA: a genetic system with eight building blocks
1
2019
... 针对核酸的碱基、磷酸和五碳糖三个主要组成部分,可以分别进行人工修饰或替换,形成非天然核酸(XNA)这样一类核酸分子类似物[219].XNA能够通过碱基修饰或骨架修饰来拓展遗传信息的存储能力,可以具有部分或全部天然核酸的属性[220].生物体系引入XNA能够拓展遗传信息的多样性,进而拓展生物性能.非天然碱基研究近年来取得了一些重大突破,如八碱基DNA和具有生命活性的六碱基DNA等.通过对天然碱基(A、T、C、G)的分子结构进行修饰,创造出了两对新的可以互补配对的碱基(S、B、P、Z,其中S和B配对、P和Z配对).将人工碱基与天然碱基相结合,可以获得含有八种碱基的DNA[221].研究者在探索非天然碱基多样性的同时,也在尝试使非天然碱基具有生物活性.美国学者在大肠杆菌里成功表达了含有两个非天然碱基编码非天然氨基酸的荧光蛋白,实现了非天然碱基的转录活性,首次实现含六碱基DNA的半人工合成生命[218].XNA种类的增加不仅提升了遗传信息的密度,还为拓展生物的多样性奠定了基础. ...
Non canonical genetic material
1
2019
... 目前XNA主要通过外源添加方式进入生物体[218],将来开发生物体内合成XNA,将进一步拓展XNA在合成生物学领域的广泛应用.由于XNA在体内的稳定性和特异性有其自身特点,在合成生物学的精准检测和正交控制等方面具有重要的应用潜力[222]. ...
Photoelectrochemical reduction of carbon dioxide to methanol through a highly efficient enzyme cascade
1
2017
... 利用生物酶与人工材料结合,形成具有部分生命特征的耦合系统是当前的一个研究热点.其中,模拟自然界光合作用,将生物酶与半导体光催化剂结合,构建基于酶-光耦联的人工光合系统发展迅速.与自然界光合作用相比,酶-光耦联系统既保留了生物酶的催化能力,又结合了半导体的光吸收能力,通过二者体外灵活组合,打破了自然进化的限制,实现太阳能驱动能源化学品和高附加值化学品的绿色、可持续合成.酸脱氢酶、醛脱氢酶和醇脱氢酶与α-Fe2O3/BiFeO3半导体光电极耦合,实现了太阳能驱动二氧化碳到甲醇还原[223].光系统(PS Ⅱ)和氢酶与半导体光电极耦合,实现了无偏压下的全水分解[224].此外,半导体与生命单元的结合还可延伸至细胞,将半导体分别沉积于原核微生物和真核微生物细胞表面,实现了微生物胞内NADPH/NADP+比例及代谢途径的调控,突破了天然微生物的生产能力上限[225,226].为减少纳米材料与细胞膜直接接触对微生物产生的破坏作用,细菌生物被膜可用于半导体大规模锚定,构建生物化学兼容的生物-无机界面[227].该体系不仅可以发展可循环和可再生的催化反应体系,并且能与胞内的氢化酶耦合实现稳定的光催化产氢[228].半导体材料与不同酶及不同细胞的兼容组合,打破了自然界进化的限制,极大促进了人工光合领域的发展. ...
Bias-free photoelectrochemical water splitting with photosystem II on a dye-sensitized photoanode wired to hydrogenase
1
2018
... 利用生物酶与人工材料结合,形成具有部分生命特征的耦合系统是当前的一个研究热点.其中,模拟自然界光合作用,将生物酶与半导体光催化剂结合,构建基于酶-光耦联的人工光合系统发展迅速.与自然界光合作用相比,酶-光耦联系统既保留了生物酶的催化能力,又结合了半导体的光吸收能力,通过二者体外灵活组合,打破了自然进化的限制,实现太阳能驱动能源化学品和高附加值化学品的绿色、可持续合成.酸脱氢酶、醛脱氢酶和醇脱氢酶与α-Fe2O3/BiFeO3半导体光电极耦合,实现了太阳能驱动二氧化碳到甲醇还原[223].光系统(PS Ⅱ)和氢酶与半导体光电极耦合,实现了无偏压下的全水分解[224].此外,半导体与生命单元的结合还可延伸至细胞,将半导体分别沉积于原核微生物和真核微生物细胞表面,实现了微生物胞内NADPH/NADP+比例及代谢途径的调控,突破了天然微生物的生产能力上限[225,226].为减少纳米材料与细胞膜直接接触对微生物产生的破坏作用,细菌生物被膜可用于半导体大规模锚定,构建生物化学兼容的生物-无机界面[227].该体系不仅可以发展可循环和可再生的催化反应体系,并且能与胞内的氢化酶耦合实现稳定的光催化产氢[228].半导体材料与不同酶及不同细胞的兼容组合,打破了自然界进化的限制,极大促进了人工光合领域的发展. ...
Self-photosensitization of nonphotosynthetic bacteria for solar-to-chemical production
1
2016
... 利用生物酶与人工材料结合,形成具有部分生命特征的耦合系统是当前的一个研究热点.其中,模拟自然界光合作用,将生物酶与半导体光催化剂结合,构建基于酶-光耦联的人工光合系统发展迅速.与自然界光合作用相比,酶-光耦联系统既保留了生物酶的催化能力,又结合了半导体的光吸收能力,通过二者体外灵活组合,打破了自然进化的限制,实现太阳能驱动能源化学品和高附加值化学品的绿色、可持续合成.酸脱氢酶、醛脱氢酶和醇脱氢酶与α-Fe2O3/BiFeO3半导体光电极耦合,实现了太阳能驱动二氧化碳到甲醇还原[223].光系统(PS Ⅱ)和氢酶与半导体光电极耦合,实现了无偏压下的全水分解[224].此外,半导体与生命单元的结合还可延伸至细胞,将半导体分别沉积于原核微生物和真核微生物细胞表面,实现了微生物胞内NADPH/NADP+比例及代谢途径的调控,突破了天然微生物的生产能力上限[225,226].为减少纳米材料与细胞膜直接接触对微生物产生的破坏作用,细菌生物被膜可用于半导体大规模锚定,构建生物化学兼容的生物-无机界面[227].该体系不仅可以发展可循环和可再生的催化反应体系,并且能与胞内的氢化酶耦合实现稳定的光催化产氢[228].半导体材料与不同酶及不同细胞的兼容组合,打破了自然界进化的限制,极大促进了人工光合领域的发展. ...
Light-driven fine chemical production in yeast biohybrids
1
2018
... 利用生物酶与人工材料结合,形成具有部分生命特征的耦合系统是当前的一个研究热点.其中,模拟自然界光合作用,将生物酶与半导体光催化剂结合,构建基于酶-光耦联的人工光合系统发展迅速.与自然界光合作用相比,酶-光耦联系统既保留了生物酶的催化能力,又结合了半导体的光吸收能力,通过二者体外灵活组合,打破了自然进化的限制,实现太阳能驱动能源化学品和高附加值化学品的绿色、可持续合成.酸脱氢酶、醛脱氢酶和醇脱氢酶与α-Fe2O3/BiFeO3半导体光电极耦合,实现了太阳能驱动二氧化碳到甲醇还原[223].光系统(PS Ⅱ)和氢酶与半导体光电极耦合,实现了无偏压下的全水分解[224].此外,半导体与生命单元的结合还可延伸至细胞,将半导体分别沉积于原核微生物和真核微生物细胞表面,实现了微生物胞内NADPH/NADP+比例及代谢途径的调控,突破了天然微生物的生产能力上限[225,226].为减少纳米材料与细胞膜直接接触对微生物产生的破坏作用,细菌生物被膜可用于半导体大规模锚定,构建生物化学兼容的生物-无机界面[227].该体系不仅可以发展可循环和可再生的催化反应体系,并且能与胞内的氢化酶耦合实现稳定的光催化产氢[228].半导体材料与不同酶及不同细胞的兼容组合,打破了自然界进化的限制,极大促进了人工光合领域的发展. ...
Programming cells for dynamic assembly of Iiorganic nano-objects with spatiotemporal control
1
2018
... 利用生物酶与人工材料结合,形成具有部分生命特征的耦合系统是当前的一个研究热点.其中,模拟自然界光合作用,将生物酶与半导体光催化剂结合,构建基于酶-光耦联的人工光合系统发展迅速.与自然界光合作用相比,酶-光耦联系统既保留了生物酶的催化能力,又结合了半导体的光吸收能力,通过二者体外灵活组合,打破了自然进化的限制,实现太阳能驱动能源化学品和高附加值化学品的绿色、可持续合成.酸脱氢酶、醛脱氢酶和醇脱氢酶与α-Fe2O3/BiFeO3半导体光电极耦合,实现了太阳能驱动二氧化碳到甲醇还原[223].光系统(PS Ⅱ)和氢酶与半导体光电极耦合,实现了无偏压下的全水分解[224].此外,半导体与生命单元的结合还可延伸至细胞,将半导体分别沉积于原核微生物和真核微生物细胞表面,实现了微生物胞内NADPH/NADP+比例及代谢途径的调控,突破了天然微生物的生产能力上限[225,226].为减少纳米材料与细胞膜直接接触对微生物产生的破坏作用,细菌生物被膜可用于半导体大规模锚定,构建生物化学兼容的生物-无机界面[227].该体系不仅可以发展可循环和可再生的催化反应体系,并且能与胞内的氢化酶耦合实现稳定的光催化产氢[228].半导体材料与不同酶及不同细胞的兼容组合,打破了自然界进化的限制,极大促进了人工光合领域的发展. ...
Immobilization of functional nano-objects in living engineered bacterial biofilms for catalytic applications
1
2019
... 利用生物酶与人工材料结合,形成具有部分生命特征的耦合系统是当前的一个研究热点.其中,模拟自然界光合作用,将生物酶与半导体光催化剂结合,构建基于酶-光耦联的人工光合系统发展迅速.与自然界光合作用相比,酶-光耦联系统既保留了生物酶的催化能力,又结合了半导体的光吸收能力,通过二者体外灵活组合,打破了自然进化的限制,实现太阳能驱动能源化学品和高附加值化学品的绿色、可持续合成.酸脱氢酶、醛脱氢酶和醇脱氢酶与α-Fe2O3/BiFeO3半导体光电极耦合,实现了太阳能驱动二氧化碳到甲醇还原[223].光系统(PS Ⅱ)和氢酶与半导体光电极耦合,实现了无偏压下的全水分解[224].此外,半导体与生命单元的结合还可延伸至细胞,将半导体分别沉积于原核微生物和真核微生物细胞表面,实现了微生物胞内NADPH/NADP+比例及代谢途径的调控,突破了天然微生物的生产能力上限[225,226].为减少纳米材料与细胞膜直接接触对微生物产生的破坏作用,细菌生物被膜可用于半导体大规模锚定,构建生物化学兼容的生物-无机界面[227].该体系不仅可以发展可循环和可再生的催化反应体系,并且能与胞内的氢化酶耦合实现稳定的光催化产氢[228].半导体材料与不同酶及不同细胞的兼容组合,打破了自然界进化的限制,极大促进了人工光合领域的发展. ...
Cell-free protein synthesis for producing ‘difficult-to-express’ proteins
1
2018
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Cell-free methods to produce structurally intact mammalian membrane proteins
1
2016
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Rapid and scalable characterization of CRISPR technologies using an E. coli cell-free transcription-translation system
1
2018
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Development of a CHO-based cell-free platform for synthesis of active monoclonal antibodies
1
2017
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Cell-free expression, purification and immunoreactivity assessment of recombinant Fasciola hepatica saposin-like protein-2
1
2018
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Advances and challenges in cell-free incorporation of unnatural amino acids into proteins
1
2019
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Production of site-specific antibody-drug conjugates using optimized non-natural amino acids in a cell-free expression system
1
2014
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Protein labeling strategies for liquid-state NMR spectroscopy using cell-free synthesis
1
2018
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Cell-free synthetic biology: engineering beyond the cell
1
2016
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...
Cell-free protein synthesis using S30 extracts from Escherichia coli RFzero strains for efficient incorporation of non-natural amino acids into proteins
1
2019
... 无细胞蛋白合成系统是在试管内重现生命中心法则的全部生化反应,可以重建自然并超越自然的生物分子系统.无细胞蛋白合成系统(cell-free)有助于实现对蛋白质表达体系的高效优化与控制,是合成理化性质复杂的蛋白质的重要方法,如生理毒性蛋白[229]、复杂膜蛋白[230]等.该系统也可以辅助对蛋白质功能的筛选与检测,如将cell-free合成系统与CRISPR-Cas系统相结合来筛选核酸酶[231].cell-free合成系统能够为活性抗体药物的表达与筛选提供便利,大大缩短活性抗体的研究周期[232].利用真核生物无细胞系统TNTT7快速体外表达FhSAP2蛋白,有效保持了其主要的构象和抗原性质[233].通过cell-free系统在天然蛋白质结构中插入非天然氨基酸也是近年来各种基础和应用科学的研究热点[234].cell-free合成系统可以为抗体偶联药物制备偶联药物前体,消除了内源性氨基酸残基随机生成的偶联物所产生的异质性和不稳定性[235].在检测方面,cell-free合成系统可以为蛋白质共振配位提供多样性的标记策略,制备用于液相核磁共振波谱分析的蛋白质样品[236].各种cell-free合成系统的开发使得廉价和快速的重组蛋白表达成为可能,并且各种方法之间能够为表达来自不同生物体的蛋白质进行替代和互补[237].在多样的cell-free合成系统中,基于大肠杆菌无细胞合成系统得到广泛的研究与应用.基于大肠杆菌RFzero-iy菌株的S30无细胞合成系统实现了高达90%以上的3-碘-L-酪氨酸的插入效率[238].鉴于系统的灵活性,无细胞蛋白质合成系统已经成为合成生物学的重要工具. ...



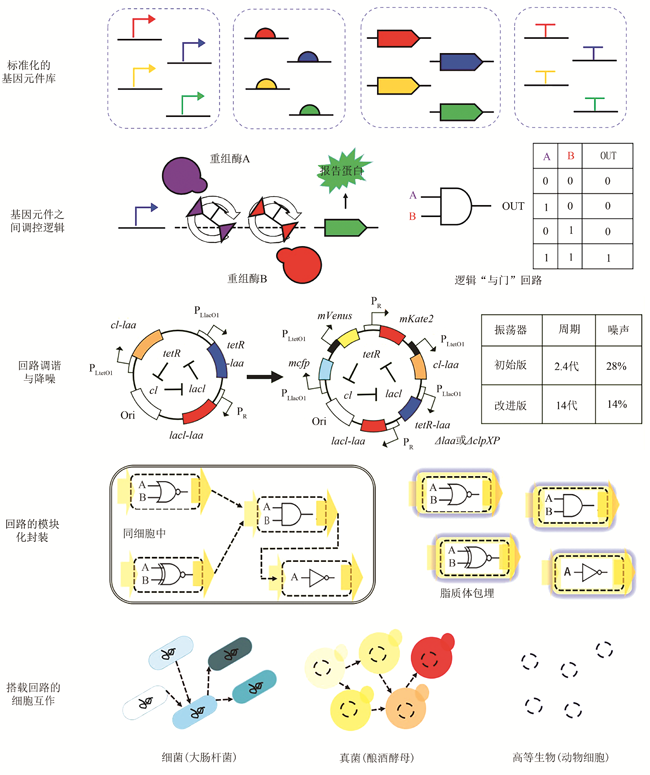
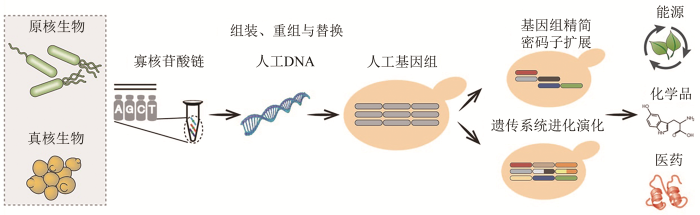
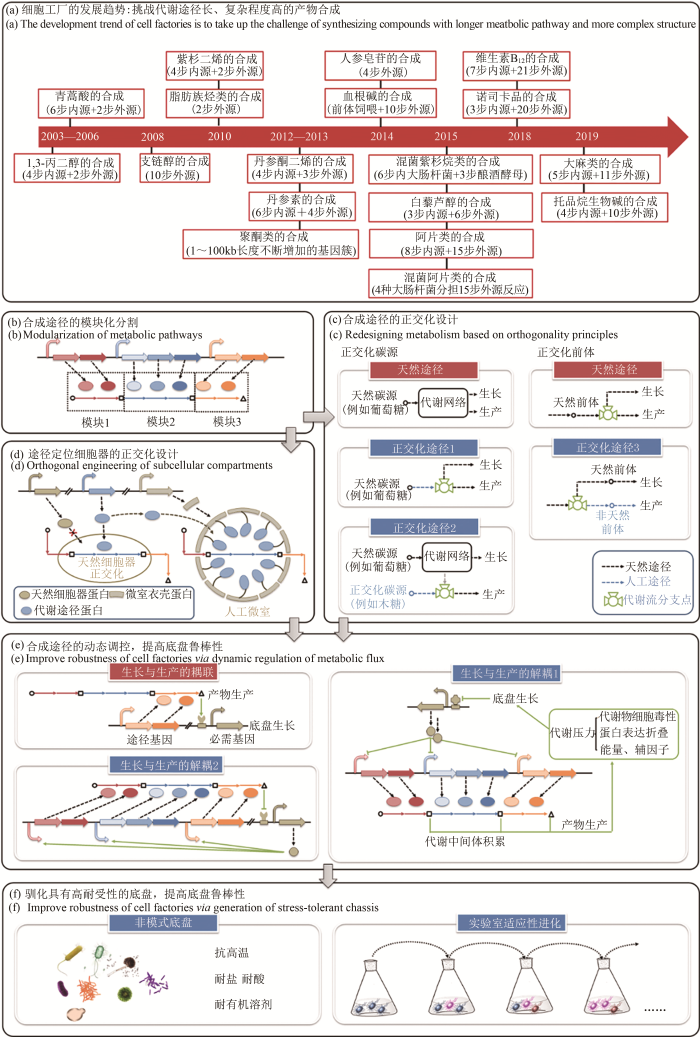
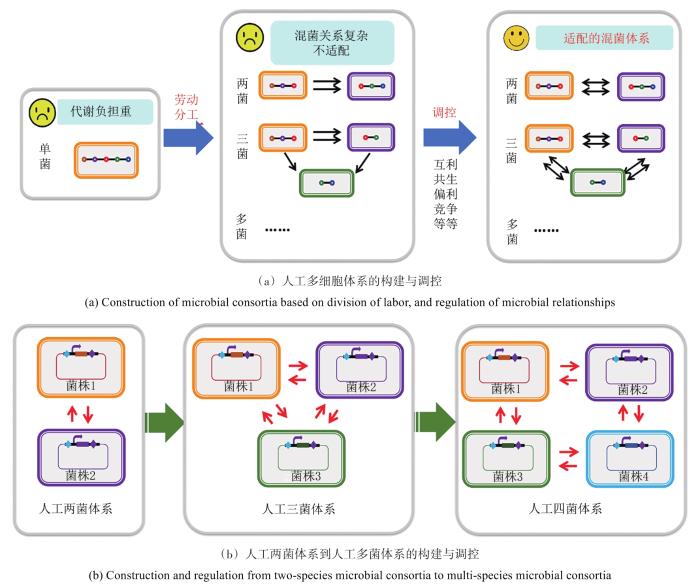


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}