|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
植物合成生物学:植物细胞大规模培养的新机遇
合成生物学
2025, 6 (5):
1107-1125.
DOI:10.12211/2096-8280.2024-095
植物细胞培养(plant cell culture, PCC)作为一种极具发展潜力的生物合成平台,具有生长周期短、成本效益高、无病原危害、次生代谢产物丰富等优势,在医药、食品和保健等领域备受关注。然而,生产效率不足是限制PCC应用于商业化生产的最大阻碍,其中,遗传转化效率低、调控网络复杂、细胞结团及遗传稳定性差是主要困难。合成生物学遵循自下而上的工程化建造理念,对天然植物细胞进行精准设计与改造,为开发高效、经济可行的植物细胞工厂提供了新的解决方案。本文回顾了PCC作为合成平台在生产重组蛋白和次生代谢产物中的研究现状。重点探讨了植物合成生物学对PCC在工业化发展中的推动作用,包括优质植物细胞系的构建、遗传转化体系的优化、表达系统的优化、生产效率与产能的提升以及赋予植物细胞合成异源产物的能力。未来,PCC的发展更需强调合成生物学理念和技术在突破当前技术瓶颈中的关键作用,以促进植物细胞大规模培养的进一步发展。
表2
植物细胞异源合成重组蛋白
正文中引用本图/表的段落
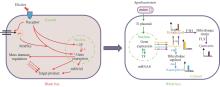
植物细胞独特的结构赋予其优越的生物制造特性。坚韧的细胞壁提供支持和保护,巨大的液泡不仅能储存潜在的毒性物质,还能调节渗透压以维持稳态,使植物细胞具备卓越的机械强度和抗逆性。此外,用于疾病治疗的功能蛋白在哺乳动物细胞中往往具有生理活性,从而影响细胞生长甚至产生细胞毒性,而植物细胞通常不受这些限制,这使其在复杂蛋白的安全、稳定、高效生产上具有显著优势。PCC生产重组蛋白技术已经被广泛用于药用蛋白(如抗原、疫苗、抗菌蛋白)、胶原蛋白和工业酶的商业化生产[13](表2)。
调控元件决定了基因的表达模式与表达强度,主要包括启动子、5′和3′非编码区、终止子、增强子、转录因子、顺式作用元件等DNA序列和蛋白序列。在植物细胞体系中,通过将调控元件序列和报告基因构建成基本表达盒或简单遗传回路,转化到植物细胞中进行定量表征,可用于优化植物细胞表达系统。例如,通过双荧光素酶报告系统(萤火虫荧光素酶/海肾荧光素酶),对不同植物启动子在BY2愈伤细胞中进行定量表征,可获得不同梯度强度的定量数据,用于精细控制蛋白表达水平和目标产物产量[85]。稳定的转基因植物细胞培养体系通常需要经过长时间的筛选过程,因此,将愈伤组织或悬浮细胞经酶解制备成原生质体是元件快速表征的一种高效替代方法。Ko Kato团队[86]通过制备水稻(O. sativa)悬浮培养细胞、黑麦(Secale cereale L.)叶片和拟南芥(A. thaliana)悬浮细胞的原生质体,在三个体系中统一验证了15个5′UTR序列的增强子活性。这种方法显著提升了元件表征效率,为调控元件在植物细胞中的应用提供了重要依据。
利用植物合成生物学能够突破植物合成能力的限制,让植物细胞得以合成原本无法产生的高附加值产物。例如,在胡萝卜愈伤组织细胞中异源表达六个拟南芥来源的苄基硫代葡萄糖苷生物合成途径基因,成功在细胞中实现了苄基硫代葡萄糖苷的积累,产量达到2.5 nmol/g FW[102]。
绿原酸1799.69 ng/mL培养液; ... Metabolic engineering of artificially modified transcription factor SmMYB36-VP16 for high-level production of tanshinones and phenolic acids 1 2024 ... 基于转录因子强大的协同调控能力,只需要调控单个转录因子就能实现对途径上多个基因的表达优化,进而特异性提高PCC中目标产物的产能.在人参愈伤组织细胞中,过表达转录因子pgNAC072能够上调人参皂苷生物合成途径中的关键酶基因表达,提高人参皂苷的积累[ Redesigning regulatory components of quorum-sensing system for diverse metabolic control 1 2022 ... 动态遗传回路通常利用底盘细胞对诱导物、温度、细胞群体或环境压力等信号的响应来控制基因的表达,这一策略在微生物底盘的成功应用充分证明了其在降低合成途径对菌体生长的影响以及提高目标产物产量上的潜力.袁其朋研究团队[ Developing a pathway-independent and full-autonomous global resource allocation strategy to dynamically switching phenotypic states 1 2020 ... 动态遗传回路通常利用底盘细胞对诱导物、温度、细胞群体或环境压力等信号的响应来控制基因的表达,这一策略在微生物底盘的成功应用充分证明了其在降低合成途径对菌体生长的影响以及提高目标产物产量上的潜力.袁其朋研究团队[ Production of benzylglucosinolate in genetically engineered carrot suspension cultures 2 2022 ... 利用植物合成生物学能够突破植物合成能力的限制,让植物细胞得以合成原本无法产生的高附加值产物.例如,在胡萝卜愈伤组织细胞中异源表达六个拟南芥来源的苄基硫代葡萄糖苷生物合成途径基因,成功在细胞中实现了苄基硫代葡萄糖苷的积累,产量达到2.5 nmol/g FW[
本文的其它图/表
|