合成生物学 ›› 2023, Vol. 4 ›› Issue (1): 225-240.DOI: 10.12211/2096-8280.2022-051
• 研究论文 • 上一篇
基因增变器驱动的酿酒酵母基因组连续进化
潘颖佳1,2, 夏思杨1, 董昌2, 蔡谨1, 连佳长1,2
- 1.浙江大学化学工程与生物工程学院,浙江 杭州 310027
2.浙江大学杭州国际科创中心,浙江 杭州 310000
-
收稿日期:2022-09-21修回日期:2022-12-14出版日期:2023-02-28发布日期:2023-03-07 -
通讯作者:蔡谨,连佳长 -
作者简介:潘颖佳 (1992—),女,博士研究生。研究方向为合成生物学。E-mail:12228048@zju.edu.cn夏思杨 (1996—),女,硕士研究生。研究方向为基因组进化。E-mail:21828174@zju.edu.cn蔡谨 (1960—),男,博士,副教授。研究方向为工业微生物学。E-mail:caij@zju.edu.cn连佳长 (1984—),男,博士,研究员。研究方向为合成生物学。E-mail:jzlian@zju.edu.cn -
基金资助:国家自然科学基金(21808199);国家重点研发计划(2018YFA0901800);浙江省杰出青年基金(LR20B060003);中央高校基本科研业务费专项资金(226-2022-00214)
Mutator-driven continuous genome evolution of Saccharomyces cerevisiae
PAN Yingjia1,2, XIA Siyang1, DONG Chang2, CAI Jin1, LIAN Jiazhang1,2
- 1.Key Laboratory of Biomass Chemical Engineering of Ministry of Education,College of Chemical and Biological Engineering,Zhejiang University,Hangzhou 310027,Zhejiang,China
2.ZJU-Hangzhou Global Scientific and Technological Innovation Center,Zhejiang University,Hangzhou 310000,Zhejiang,China
-
Received:2022-09-21Revised:2022-12-14Online:2023-02-28Published:2023-03-07 -
Contact:CAI Jin, LIAN Jiazhang
摘要:
酿酒酵母是工业生物技术最常用的底盘细胞之一,被广泛应用于生物基化学品和高附加值产品的大规模生产。鉴于生物体系代谢和调控网络的复杂性,由多基因协同控制的复杂生理性状的改造通常需要采取全基因组进化来实现。为实现酿酒酵母基因组的快速进化,本研究采用CRISPR干扰技术(CRISPRi)调控与染色体复制和稳定性相关基因(MSH2、TSA1、RAD27和CLB5,即增变基因)的表达水平,构建了能够调控酿酒酵母基因组突变率和突变类型的基因增变器。利用基因增变器提高酿酒酵母的β-胡萝卜素合成水平、木糖利用效率和异丁醇耐受性。此外,构建了混合基因增变器和多重基因增变器,进一步探究了不同表型基因组进化所需的最佳突变类型以及不同突变类型之间的协同进化机制。本研究不仅可用于创建高性能酿酒酵母细胞工厂,还有可能发展一个具有普遍适用性的基因组连续进化策略。
中图分类号:
引用本文
潘颖佳, 夏思杨, 董昌, 蔡谨, 连佳长. 基因增变器驱动的酿酒酵母基因组连续进化[J]. 合成生物学, 2023, 4(1): 225-240.
PAN Yingjia, XIA Siyang, DONG Chang, CAI Jin, LIAN Jiazhang. Mutator-driven continuous genome evolution of Saccharomyces cerevisiae[J]. Synthetic Biology Journal, 2023, 4(1): 225-240.
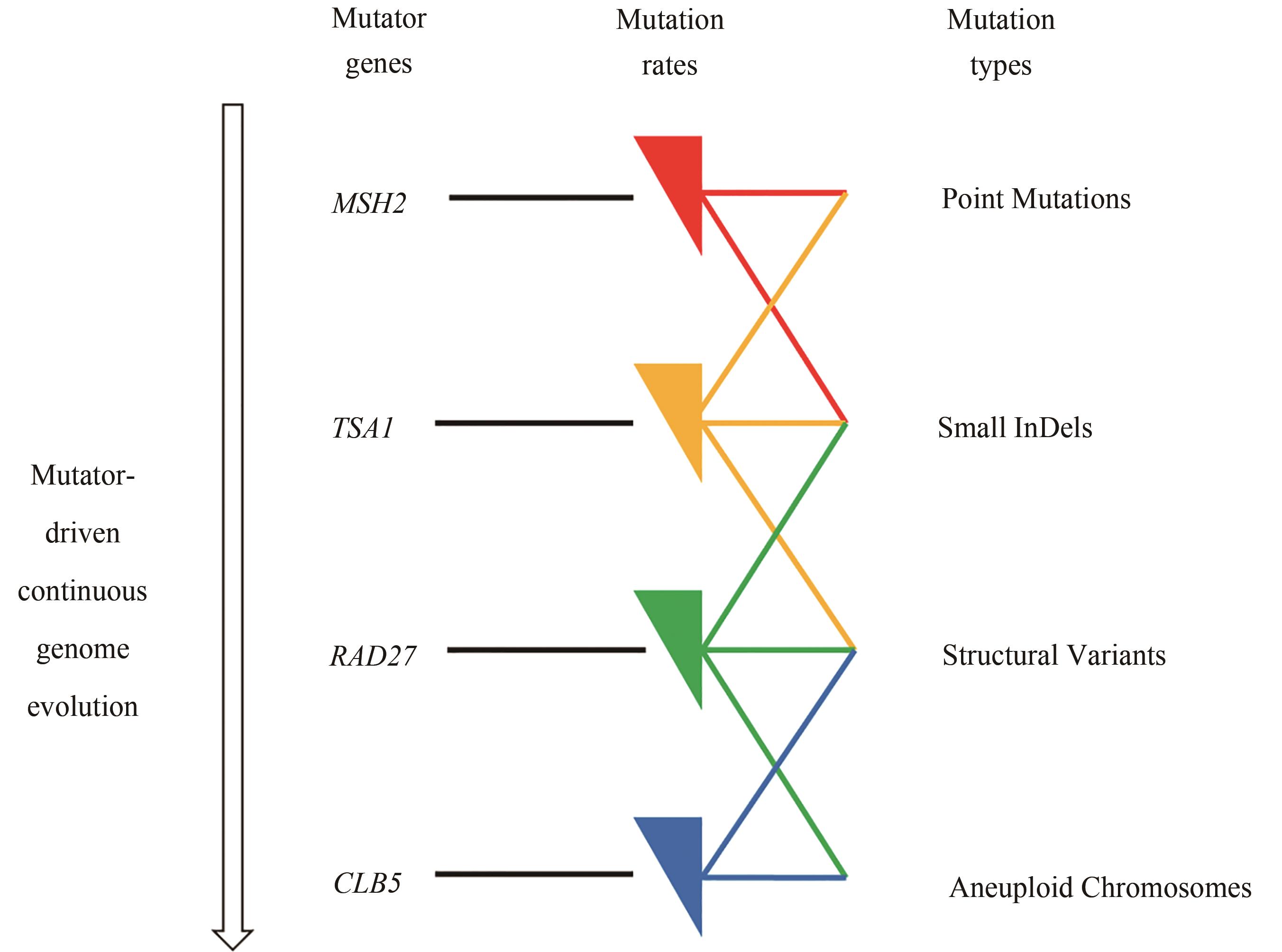
图1 基于增变基因的酿酒酵母基因组连续进化[酿酒酵母中存在一些维持遗传稳定性的基因,其敲除或抑制会引发不同的突变类型,提高基因组突变率,即增变基因(mutator genes)。利用CRISPRi系统抑制增变基因的表达水平,构建4个基因增变器(mutators),可诱导酿酒酵母基因组发生不同类型的突变]
Fig. 1 Continuous genome evolution of S. cerevisiae through mutator genes(There are mutator genes in S. cerevisiae to maintain its genetic stability, and their knockdown or suppression can increase the genome mutation rate, and also result in different types of mutations. Using the CRISPRi system to inhibit the expression of mutator genes, four mutators were constructed to induce different types of mutations in the genome of S. cerevisiae.)
| Strains | Description | Source |
|---|---|---|
| BY4741-iAID6 | BY4741 with CRISPRi machinery (dSpCas9-RD1152) integrated | Our lab[ |
| y426 | BY4741-iAID6/p426 | This study |
| yMM | BY4741-iAID6/MM | This study |
| yTM | BY4741-iAID6/TM | This study |
| yRM | BY4741-iAID6/RM | This study |
| yCM | BY4741-iAID6/CM | This study |
| BY4741-iAID6-CrtIEYB | BY4741-iAID6-CrtE-CrtYB-CrtI | Our lab[ |
| yCar426 | BY4741-iAID6-CrtE-CrtYB-CrtI/p426 | This study |
| yCarMM | BY4741-iAID6-CrtE-CrtYB-CrtI/MM | This study |
| yCarTM | BY4741-iAID6-CrtE-CrtYB-CrtI/TM | This study |
| yCarRM | BY4741-iAID6-CrtE-CrtYB-CrtI/RM | This study |
| yCarCM | BY4741-iAID6-CrtE-CrtYB-CrtI/CM | This study |
| BY4741-iAID6-psXP | BY4741-iAID6-XR-XDH-XKS | This study |
| yX426 | BY4741-iAID6-XR-XDH-XKS/p426 | This study |
| yXMM | BY4741-iAID6-XR-XDH-XKS/MM | This study |
| yXRM | BY4741-iAID6-XR-XDH-XKS/RM | This study |
| yXTM | BY4741-iAID6-XR-XDH-XKS/TM | This study |
| yXCM | BY4741-iAID6-XR-XDH-XKS/CM | This study |
| yXMTRC | BY4741-iAID6-XR-XDH-XKS/MTRC | This study |
| yXMTRC*2 | BY4741-iAID6-XR-XDH-XKS/MTRC-MTRC | This study |
| CT | CEN-PK2-1C; TEF1p-mVenus-PGK1t; URA3 | Our lab[ |
| MSH2p-mVenus | BY4741-iAID6/p415-MSH2p-mVenus | This study |
| TSA1p-mVenus | BY4741-iAID6/p415-TSA1p-mVenus | This study |
| RAD27p-mVenus | BY4741-iAID6/p415-RAD27p-mVenus | This study |
| CLB5p-mVenus | BY4741-iAID6/p415-CLB5p-mVenus | This study |
表1 本研究相关的酿酒酵母菌株
Table 1 S. cerevisiae strains used in this study
| Strains | Description | Source |
|---|---|---|
| BY4741-iAID6 | BY4741 with CRISPRi machinery (dSpCas9-RD1152) integrated | Our lab[ |
| y426 | BY4741-iAID6/p426 | This study |
| yMM | BY4741-iAID6/MM | This study |
| yTM | BY4741-iAID6/TM | This study |
| yRM | BY4741-iAID6/RM | This study |
| yCM | BY4741-iAID6/CM | This study |
| BY4741-iAID6-CrtIEYB | BY4741-iAID6-CrtE-CrtYB-CrtI | Our lab[ |
| yCar426 | BY4741-iAID6-CrtE-CrtYB-CrtI/p426 | This study |
| yCarMM | BY4741-iAID6-CrtE-CrtYB-CrtI/MM | This study |
| yCarTM | BY4741-iAID6-CrtE-CrtYB-CrtI/TM | This study |
| yCarRM | BY4741-iAID6-CrtE-CrtYB-CrtI/RM | This study |
| yCarCM | BY4741-iAID6-CrtE-CrtYB-CrtI/CM | This study |
| BY4741-iAID6-psXP | BY4741-iAID6-XR-XDH-XKS | This study |
| yX426 | BY4741-iAID6-XR-XDH-XKS/p426 | This study |
| yXMM | BY4741-iAID6-XR-XDH-XKS/MM | This study |
| yXRM | BY4741-iAID6-XR-XDH-XKS/RM | This study |
| yXTM | BY4741-iAID6-XR-XDH-XKS/TM | This study |
| yXCM | BY4741-iAID6-XR-XDH-XKS/CM | This study |
| yXMTRC | BY4741-iAID6-XR-XDH-XKS/MTRC | This study |
| yXMTRC*2 | BY4741-iAID6-XR-XDH-XKS/MTRC-MTRC | This study |
| CT | CEN-PK2-1C; TEF1p-mVenus-PGK1t; URA3 | Our lab[ |
| MSH2p-mVenus | BY4741-iAID6/p415-MSH2p-mVenus | This study |
| TSA1p-mVenus | BY4741-iAID6/p415-TSA1p-mVenus | This study |
| RAD27p-mVenus | BY4741-iAID6/p415-RAD27p-mVenus | This study |
| CLB5p-mVenus | BY4741-iAID6/p415-CLB5p-mVenus | This study |
| Mutators | gRNA plasmids | ||||
|---|---|---|---|---|---|
| MM | p426-Sg11 | p426-Sg12 | p426-Sg15 | p426-Sg17 | p426-SpSgH |
| TM | p426-Sg21 | p426-Sg22 | p426-Sg24 | p426-Sg26 | p426-SpSgH |
| RM | p426-Sg32 | p426-Sg33 | p426-Sg34 | p426-Sg36 | p426-SpSgH |
| CM | p426-Sg41 | p426-Sg42 | p426-Sg44 | p426-Sg45 | p426-SpSgH |
表2 不同基因增变器对应的gRNA 质粒文库
Table 2 gRNA plasmid libraries for mutators
| Mutators | gRNA plasmids | ||||
|---|---|---|---|---|---|
| MM | p426-Sg11 | p426-Sg12 | p426-Sg15 | p426-Sg17 | p426-SpSgH |
| TM | p426-Sg21 | p426-Sg22 | p426-Sg24 | p426-Sg26 | p426-SpSgH |
| RM | p426-Sg32 | p426-Sg33 | p426-Sg34 | p426-Sg36 | p426-SpSgH |
| CM | p426-Sg41 | p426-Sg42 | p426-Sg44 | p426-Sg45 | p426-SpSgH |
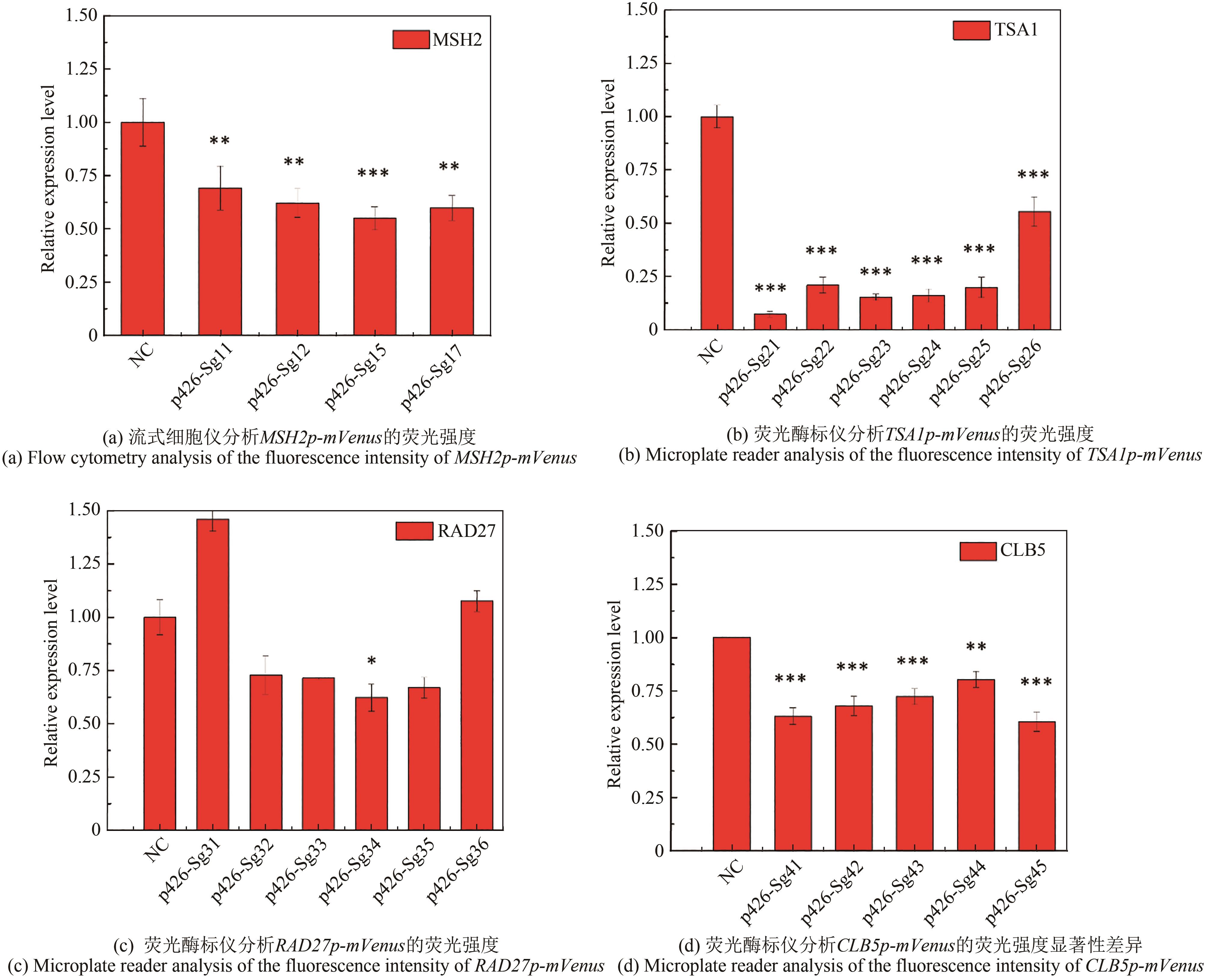
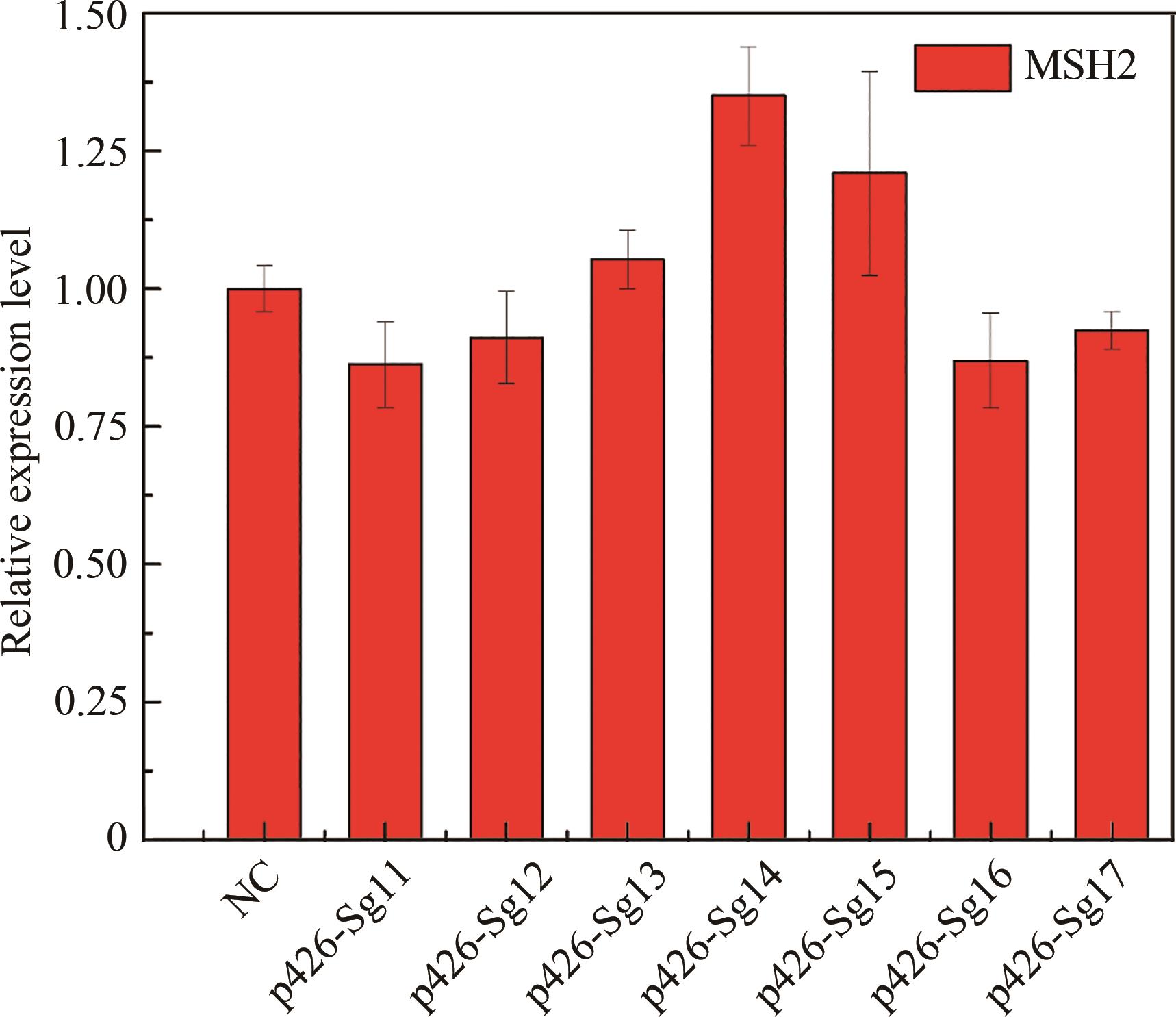
图2 以mVenus为报告基因检测不同gRNA对增变基因表达的抑制效果(显著性差异:*表示p < 0.05;**表示p < 0.01;***表示p < 0.001)
Fig. 2 Inhibition efficiency of different gRNAs on the expression of mutator genes with mVenus as a reporter(*, ** and *** for the significance p < 0.05, 0.01 and 0.001, respectively)

图3 基因增变器诱导酿酒酵母基因组产生的突变率和突变类型(a)基因增变器诱导酿酒酵母基因组产生不同的相对突变率;(b)基因增变器均诱导酿酒酵母基因组产生不同的突变类型,4个基因增变器均产生了点突变,其中TM和RM诱导产生短片段缺失和插入,RM和CM还诱导产生了中长片段插入缺失等其他突变类型(显著性差异:*表示p < 0.05;**表示p < 0.01)
Fig. 3 Mutation rates and types in the genome of S. cerevisiae induced by mutators(a) Relative mutation rates of the genome of S. cerevisiae induced by mutators (* for significance p < 0.05); (b) Mutation type of the genome of S. cerevisiae induced by mutators. All 4 mutators induced SNPs, TM and RM induced small Indels, and RM and CM also induced other mutation types. * and ** for significance p < 0.05 and 0.01, respectively.
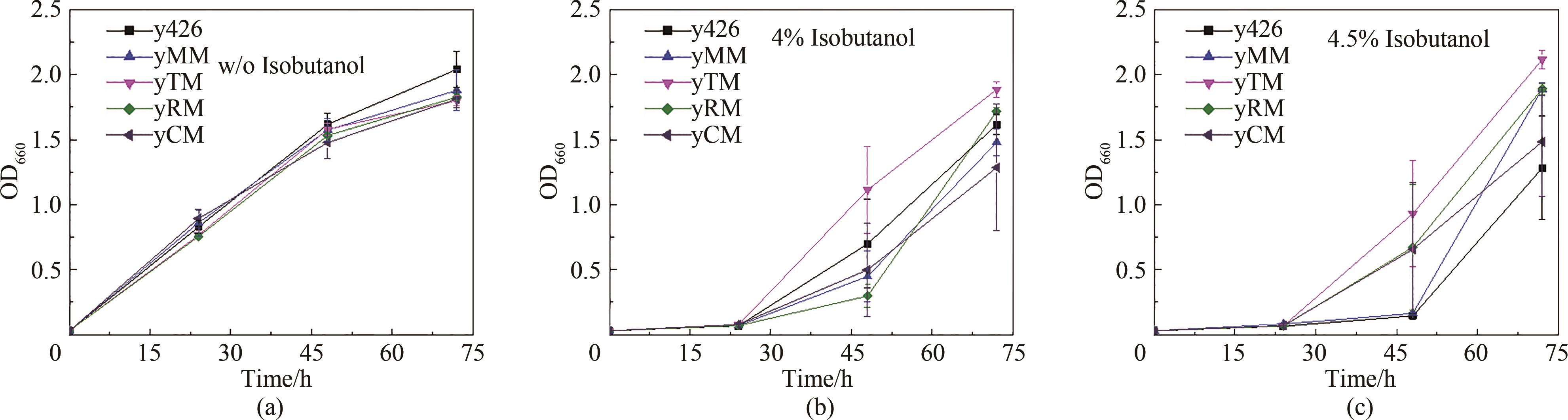
图4 基因增变器提高酿酒酵母异丁醇耐受性(a)当培养基中不存在异丁醇时,进化组和对照组之间的生长趋势无显著差异;(b)当培养基中异丁醇浓度达到4%时,yTM的生长速率显著高于其他菌株;(c)当异丁醇浓度达到4.5%时,相比对照y426菌株,各进化组都具有显著的生长优势,其中提升效果最好的增变酵母为yTM
Fig. 4 Evaluation of the mutators for improved isobutanol tolerance(a) No significant difference in the growth profiles of the mutated and control strains when isobutanol was not present in the medium; (b) When the concentration of isobutanol in the medium reached 4%, the growth rate of yTM was significantly higher than that of other strains; (c) When isobutanol concentration reached 4.5%, all mutated strains showed significant growth advantages over the control, with yTM as the best strain.
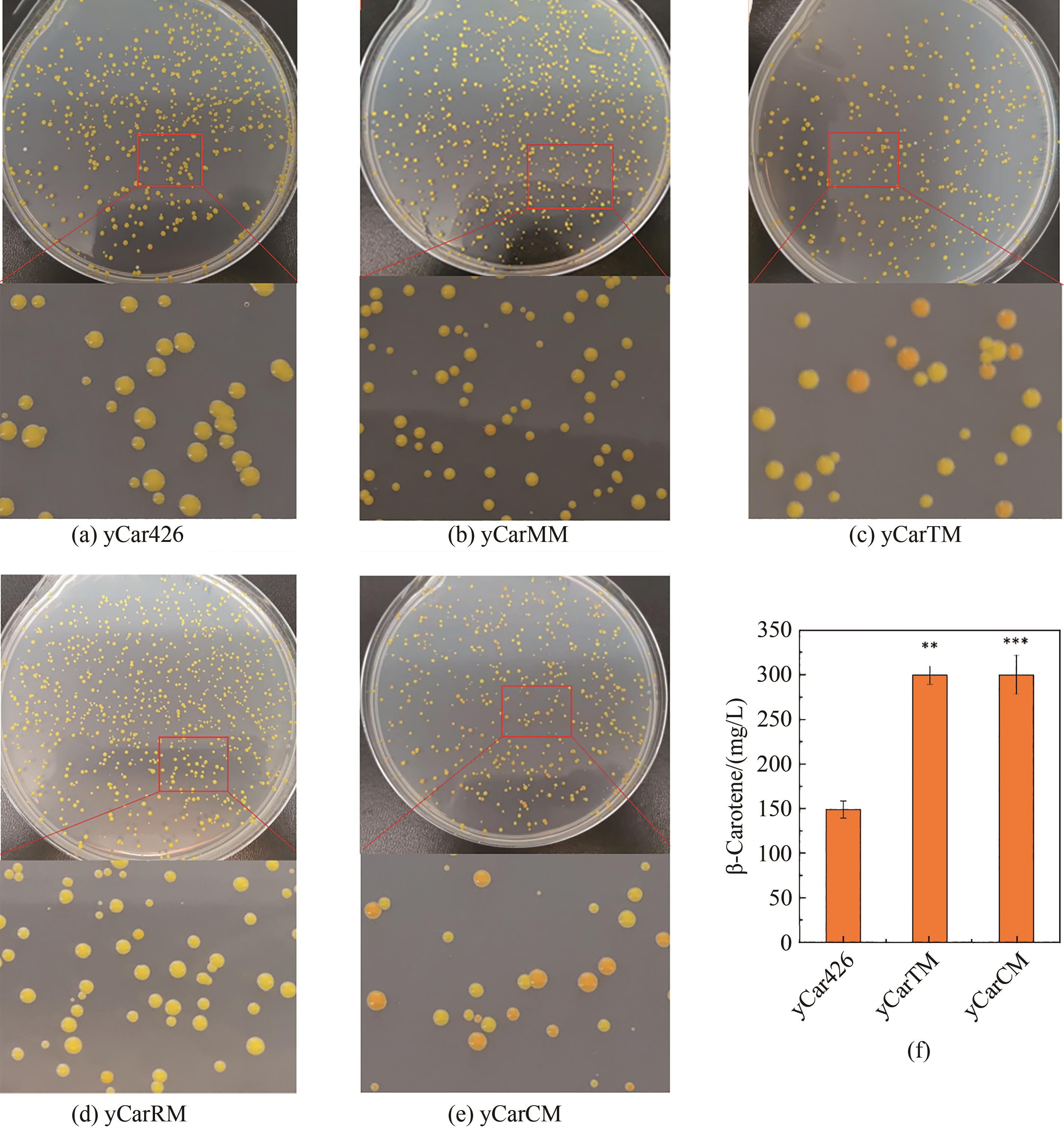
图5 基因增变器提高酿酒酵母β-胡萝卜素产量(a)~(e)基因增变器诱导酿酒酵母产生颜色深浅不一的菌落;(f)通过HPLC测定进化菌株的β-胡萝卜素产量(显著性差异:**表示p < 0.01;***表示p < 0.001)
Fig. 5 Evaluation of the mutators in increasing β-carotene biosynthesis(a)~(e) Mutation resulted in the formation of colonies with different color densities; (f) Production of β-carotene in the mutated and control strains determined by HPLC. ** and *** for significance p < 0.01 and 0.001, respectively.

图6 通过基因增变器提高酿酒酵母对木糖的利用率[不同基因增变器诱导的酿酒酵母在以木糖为唯一碳源的液体培养基中的生长曲线(a)和木糖消耗曲线(b);(c)混合增变器MTRC和多重增变器MTRC*2诱导的酿酒酵母的生长曲线(空心图例)和木糖消耗曲线(实心图例);(d)混合增变器MTRC诱导酿酒酵母在以木糖为唯一碳源的液体培养基中连续传代培养后菌液中的gRNA丰度分布]
Fig. 6 Evaluation of the mutators in improving xylose utilization[Profiles for growth (a) and xylose consumption (b) of the mutated and control strains with xylose as the sole carbon source. (c) Profiles for growth (hollow symbols) and xylose consumption (solid symbol) of the single (MTRC) and double (MTRC*2) mutated strains. (d) Distribution of gRNA abundance when the MTRC mutated strain was continuously subcultured in the liquid medium containing xylose.]
附图1 荧光酶标仪分析MSH2p-mVenus的CRISPRi结果Supplementary Fig. S1 Repression of MSH2p-mVenus by CRISPRi analyzed by a microplate reader

附图2 产β-胡萝卜素的酿酒酵母进化菌株的遗传稳定性(eCarTM菌株丢质粒前所携带的质粒为p426-Sg25,eCarCM菌株丢质粒前所携带的质粒为p426-Sg41,为验证菌株获得目的性状后能够保持遗传性状的稳定性,将获得目的性状的进化菌株去除其gRNA质粒,然后再对其性状的稳定性进行测试。以产β-胡萝卜素的进化菌株为例,具体方法如下:①在增变菌株yCarMM、yCarTM、yCarRM、yCarCM和对照菌株yCar426生长的SED-URA/G418 固体培养基上挑取颜色最深的菌落,每类菌种各挑3个单菌落,接种于SED完全液体培养基中,连续传代培养2次,同时通过测序确认所挑取的单菌落的gRNA种类;②取5 μL菌液在SED完全固体培养基上划线培养,培养2~3天后,从划线的固体培养基上随机挑取16个单菌落,每个菌落都同时在SED-URA/G418和 SED完全固体培养基上接种,筛选出只能够在SED完全固体培养基上生长的菌落,即丢弃质粒的进化菌株;③将去除质粒的进化菌株继续在SED完全液体培养基中连续传代培养3~5次后,取菌液涂布至SED完全固体培养基,通过其菌落颜色的均一性判断性状遗传的稳定性。TM和CM的进化菌株经过传代后,其菌落颜色保持均一,证明其遗传稳定性。)Supplementary Fig. S2 Genetic stability of the evolved β-carotene producing strains
| Plasmid | Genotype | Source |
|---|---|---|
| pRS415 | CEN/ARS; Amp; LEU2 | Our lab |
| pRS406-RV500 | Amp; URA3; mVenus; mCherry | Our lab |
| p415-MSH2p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p415-TSA1p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p415-RAD27p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p415-CLB5p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p426-SpSgH | 2 μ; Amp; URA3 | Our lab |
| p426-Sg11 | p426-SpSgH target on MSH2 | This study |
| p426-Sg12 | p426-SpSgH target on MSH2 | This study |
| p426-Sg13 | p426-SpSgH target on MSH2 | This study |
| p426-Sg14 | p426-SpSgH target on MSH2 | This study |
| p426-Sg15 | p426-SpSgH target on MSH2 | This study |
| p426-Sg16 | p426-SpSgH target on MSH2 | This study |
| p426-Sg17 | p426-SpSgH target on MSH2 | This study |
| p426-Sg18 | p426-SpSgH target on MSH2 | This study |
| p426-Sg21 | p426-SpSgH target on TSA1 | This study |
| p426-Sg22 | p426-SpSgH target on TSA1 | This study |
| p426-Sg23 | p426-SpSgH target on TSA1 | This study |
| p426-Sg24 | p426-SpSgH target on TSA1 | This study |
| p426-Sg25 | p426-SpSgH target on TSA1 | This study |
| p426-Sg26 | p426-SpSgH target on TSA1 | This study |
| p426-Sg31 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg32 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg33 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg34 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg35 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg36 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg41 | p426-SpSgH target on CLB5 | This study |
| p426-Sg42 | p426-SpSgH target on CLB5 | This study |
| p426-Sg43 | p426-SpSgH target on CLB5 | This study |
| p426-Sg44 | p426-SpSgH target on CLB5 | This study |
| p426-Sg45 | p426-SpSgH target on CLB5 | This study |
附表 1 本研究所用的质粒
Supplementary Table S1 Plasmids used in this study
| Plasmid | Genotype | Source |
|---|---|---|
| pRS415 | CEN/ARS; Amp; LEU2 | Our lab |
| pRS406-RV500 | Amp; URA3; mVenus; mCherry | Our lab |
| p415-MSH2p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p415-TSA1p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p415-RAD27p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p415-CLB5p-mVenus | CEN/ARS; Amp; URA3; mVenus | This study |
| p426-SpSgH | 2 μ; Amp; URA3 | Our lab |
| p426-Sg11 | p426-SpSgH target on MSH2 | This study |
| p426-Sg12 | p426-SpSgH target on MSH2 | This study |
| p426-Sg13 | p426-SpSgH target on MSH2 | This study |
| p426-Sg14 | p426-SpSgH target on MSH2 | This study |
| p426-Sg15 | p426-SpSgH target on MSH2 | This study |
| p426-Sg16 | p426-SpSgH target on MSH2 | This study |
| p426-Sg17 | p426-SpSgH target on MSH2 | This study |
| p426-Sg18 | p426-SpSgH target on MSH2 | This study |
| p426-Sg21 | p426-SpSgH target on TSA1 | This study |
| p426-Sg22 | p426-SpSgH target on TSA1 | This study |
| p426-Sg23 | p426-SpSgH target on TSA1 | This study |
| p426-Sg24 | p426-SpSgH target on TSA1 | This study |
| p426-Sg25 | p426-SpSgH target on TSA1 | This study |
| p426-Sg26 | p426-SpSgH target on TSA1 | This study |
| p426-Sg31 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg32 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg33 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg34 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg35 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg36 | p426-SpSgH target on TRAD27 | This study |
| p426-Sg41 | p426-SpSgH target on CLB5 | This study |
| p426-Sg42 | p426-SpSgH target on CLB5 | This study |
| p426-Sg43 | p426-SpSgH target on CLB5 | This study |
| p426-Sg44 | p426-SpSgH target on CLB5 | This study |
| p426-Sg45 | p426-SpSgH target on CLB5 | This study |
| Oligo | Sequences(5′→3′) | Applications |
|---|---|---|
| SpSg11-for | gatcaatatactgaaaataaaaag | To construct gRNA plasmids targeting MSH2 |
| SpSg11-rev | aaacctttttattttcagtatatt | |
| SpSg12-for | gatcgaattttagctctggcctag | |
| SpSg12-rev | aaacctaggccagagctaaaattc | |
| SpSg13-for | gatcgcctttatccactaatctaa | |
| SpSg13-rev | aaacttagattagtggataaaggc | |
| SpSg14-for | gatcggaagttctttgccgttaca | |
| SpSg14-rev | aaactgtaacggcaaagaacttcc | |
| SpSg15-for | gatctatgtatatatagaatatta | |
| SpSg15-rev | aaactaatattctatatatacata | |
| SpSg16-for | gatcttaaaagtatgtcctccact | |
| SpSg16-rev | aaacagtggaggacatacttttaa | |
| SpSg17-for | gatcaaaattctctgatgtatcag | |
| SpSg17-rev | aaacctgatacatcagagaatttt | |
| SpSg18-for | gatcacttctataagaagtataca | |
| SpSg18-rev | aaactgtatacttcttatagaagt | |
| SpSg21-for | gatcaagtcggctggcaacaaacc | To construct gRNA plasmids targeting TSA1 |
| SpSg21-rev | aaacggtttgttgccagccgactt | |
| SpSg22-for | gatcaccaggacatatataaaggg | |
| SpSg22-rev | aaacccctttatatatgtcctggt | |
| SpSg23-for | gatcaaggcccgttgagaacggtt | |
| SpSg23-rev | aaacaaccgttctcaacgggcctt | |
| SpSg24-for | gatcaggggaaggcccgttgagaa | |
| SpSg24-rev | aaacttctcaacgggccttcccct | |
| SpSg25-for | gatcggttgtgagcaattgaacga | |
| SpSg25-rev | aaactcgttcaattgctcacaacc | |
| SpSg26-for | gatcacatacacatacatacacaa | |
| SpSg26-rev | aaacttgtgtatgtatgtgtatgt | |
| SpSg31-for | gatcaaagctttaaacgcgttagg | To construct gRNA plasmids targeting RAD27 |
| SpSg31-rev | aaaccctaacgcgtttaaagcttt | |
| SpSg32-for | gatcgcgtaacatcgcgcaaatga | |
| SpSg32-rev | aaactcatttgcgcgatgttacgc | |
| SpSg33-for | gatcaaagcgttgacagcatacat | |
| SpSg33-rev | aaacatgtatgctgtcaacgcttt | |
| SpSg34-for | gatcaagaaataggaaacggacac | |
| SpSg34-rev | aaacgtgtccgtttcctatttctt | |
| SpSg35-for | gatcgacaccggaagaaaaaatat | |
| SpSg35-rev | aaacatattttttcttccggtgtc | |
| SpSg36-for | gatcaggtttgaatgcaattatat | |
| SpSg36-rev | aaacatataattgcattcaaacct | |
| SpSg41-for | gatcaattggccgtgaaaagcttt | To construct gRNA plasmids targeting CLB5 |
| SpSg41-rev | aaacaaagcttttcacggccaatt | |
| SpSg42-for | gatcctttgtgtgagacaactaat | |
| SpSg42-rev | aaacattagttgtctcacacaaag | |
| SpSg43-for | gatcgttcagcggctttaaataca | |
| SpSg43-rev | aaactgtatttaaagccgctgaac | |
| SpSg44-for | gatctgaacacctttactgaacaa | |
| SpSg44-rev | aaacttgttcagtaaaggtgttca | |
| SpSg45-for | gatctaatactctgctcatggtcg | |
| SpSg45-rev | aaaccgaccatgagcagagtatta | |
| MSH2p-for | NNNNNctcgagattagaattaaaatgtgtag | Used to amplify MSH2p |
| MSH2p-rev | NNNNNggatcctctctcctctgatacatcag | |
| mVenus-for | NNNNNggatccggcggcagcggcggcagcatggaattcgtgagcaagg | Used to amplify mVenus |
| mVenus-rev | NNNNNgagctccaggaagaatacac | |
| TSA1p-for | NNNNNctcgagacagcaggaaaacgaagatg | Used to amplify TSA1p |
| TSA1p-rev | NNNNNggatccgacggcagttttcttaaaag | |
| RAD27p-for | NNNNNctcgagaaacaaaaagaacagggaaag | Used to amplify RAD27p |
| RAD27p-rev | NNNNNggatccagcagagggaacatgttccg | |
| CLB5p-for | NNNNNctcgagtgaagacgcgcccttgatgg | Used to amplify CLB5p |
| CLB5p-rev | NNNNNggatccaatcatagaatttcttttaatac | |
| SpSg-GJ-for | cttcctgggtctcagtcccctcgagcacagggtaataact | To construct gRNA plasmid library MTRC*2 |
| SpSg-GJ-rev | tatctctaggtctcagtgggagctccctgcaggcatg | |
| 2gRNA-F1 | NNNNNggtctccggactctttgaaaagataatgtatg | |
| 2gRNA-R1 | NNNNNggtctcccggacttgcatgcctgcagggagctc | |
| 2gRNA-F2 | NNNNNggtctcctccgtctttgaaaagataatgtatg | |
| 2gRNA-R2 | NNNNNggtctcccaaccttgcatgcctgcagggagctc |
附表2 本研究所用的引物
Supplementary Table S2 Primers used in this study
| Oligo | Sequences(5′→3′) | Applications |
|---|---|---|
| SpSg11-for | gatcaatatactgaaaataaaaag | To construct gRNA plasmids targeting MSH2 |
| SpSg11-rev | aaacctttttattttcagtatatt | |
| SpSg12-for | gatcgaattttagctctggcctag | |
| SpSg12-rev | aaacctaggccagagctaaaattc | |
| SpSg13-for | gatcgcctttatccactaatctaa | |
| SpSg13-rev | aaacttagattagtggataaaggc | |
| SpSg14-for | gatcggaagttctttgccgttaca | |
| SpSg14-rev | aaactgtaacggcaaagaacttcc | |
| SpSg15-for | gatctatgtatatatagaatatta | |
| SpSg15-rev | aaactaatattctatatatacata | |
| SpSg16-for | gatcttaaaagtatgtcctccact | |
| SpSg16-rev | aaacagtggaggacatacttttaa | |
| SpSg17-for | gatcaaaattctctgatgtatcag | |
| SpSg17-rev | aaacctgatacatcagagaatttt | |
| SpSg18-for | gatcacttctataagaagtataca | |
| SpSg18-rev | aaactgtatacttcttatagaagt | |
| SpSg21-for | gatcaagtcggctggcaacaaacc | To construct gRNA plasmids targeting TSA1 |
| SpSg21-rev | aaacggtttgttgccagccgactt | |
| SpSg22-for | gatcaccaggacatatataaaggg | |
| SpSg22-rev | aaacccctttatatatgtcctggt | |
| SpSg23-for | gatcaaggcccgttgagaacggtt | |
| SpSg23-rev | aaacaaccgttctcaacgggcctt | |
| SpSg24-for | gatcaggggaaggcccgttgagaa | |
| SpSg24-rev | aaacttctcaacgggccttcccct | |
| SpSg25-for | gatcggttgtgagcaattgaacga | |
| SpSg25-rev | aaactcgttcaattgctcacaacc | |
| SpSg26-for | gatcacatacacatacatacacaa | |
| SpSg26-rev | aaacttgtgtatgtatgtgtatgt | |
| SpSg31-for | gatcaaagctttaaacgcgttagg | To construct gRNA plasmids targeting RAD27 |
| SpSg31-rev | aaaccctaacgcgtttaaagcttt | |
| SpSg32-for | gatcgcgtaacatcgcgcaaatga | |
| SpSg32-rev | aaactcatttgcgcgatgttacgc | |
| SpSg33-for | gatcaaagcgttgacagcatacat | |
| SpSg33-rev | aaacatgtatgctgtcaacgcttt | |
| SpSg34-for | gatcaagaaataggaaacggacac | |
| SpSg34-rev | aaacgtgtccgtttcctatttctt | |
| SpSg35-for | gatcgacaccggaagaaaaaatat | |
| SpSg35-rev | aaacatattttttcttccggtgtc | |
| SpSg36-for | gatcaggtttgaatgcaattatat | |
| SpSg36-rev | aaacatataattgcattcaaacct | |
| SpSg41-for | gatcaattggccgtgaaaagcttt | To construct gRNA plasmids targeting CLB5 |
| SpSg41-rev | aaacaaagcttttcacggccaatt | |
| SpSg42-for | gatcctttgtgtgagacaactaat | |
| SpSg42-rev | aaacattagttgtctcacacaaag | |
| SpSg43-for | gatcgttcagcggctttaaataca | |
| SpSg43-rev | aaactgtatttaaagccgctgaac | |
| SpSg44-for | gatctgaacacctttactgaacaa | |
| SpSg44-rev | aaacttgttcagtaaaggtgttca | |
| SpSg45-for | gatctaatactctgctcatggtcg | |
| SpSg45-rev | aaaccgaccatgagcagagtatta | |
| MSH2p-for | NNNNNctcgagattagaattaaaatgtgtag | Used to amplify MSH2p |
| MSH2p-rev | NNNNNggatcctctctcctctgatacatcag | |
| mVenus-for | NNNNNggatccggcggcagcggcggcagcatggaattcgtgagcaagg | Used to amplify mVenus |
| mVenus-rev | NNNNNgagctccaggaagaatacac | |
| TSA1p-for | NNNNNctcgagacagcaggaaaacgaagatg | Used to amplify TSA1p |
| TSA1p-rev | NNNNNggatccgacggcagttttcttaaaag | |
| RAD27p-for | NNNNNctcgagaaacaaaaagaacagggaaag | Used to amplify RAD27p |
| RAD27p-rev | NNNNNggatccagcagagggaacatgttccg | |
| CLB5p-for | NNNNNctcgagtgaagacgcgcccttgatgg | Used to amplify CLB5p |
| CLB5p-rev | NNNNNggatccaatcatagaatttcttttaatac | |
| SpSg-GJ-for | cttcctgggtctcagtcccctcgagcacagggtaataact | To construct gRNA plasmid library MTRC*2 |
| SpSg-GJ-rev | tatctctaggtctcagtgggagctccctgcaggcatg | |
| 2gRNA-F1 | NNNNNggtctccggactctttgaaaagataatgtatg | |
| 2gRNA-R1 | NNNNNggtctcccggacttgcatgcctgcagggagctc | |
| 2gRNA-F2 | NNNNNggtctcctccgtctttgaaaagataatgtatg | |
| 2gRNA-R2 | NNNNNggtctcccaaccttgcatgcctgcagggagctc |
| Strains | SNPs | Small InDels | Other mutation types |
|---|---|---|---|
| yMM | 16 | 0 | 0 |
| yTM | 12 | 4 | 0 |
| yRM | 12 | 2 | 2 |
| yCM | 4 | 0 | 12 |
附表3 基因增变器诱导酿酒酵母基因组产生的突变类型
Supplementary Table S3 Mutation types of the S. cerevisiae genome induced by mutators
| Strains | SNPs | Small InDels | Other mutation types |
|---|---|---|---|
| yMM | 16 | 0 | 0 |
| yTM | 12 | 4 | 0 |
| yRM | 12 | 2 | 2 |
| yCM | 4 | 0 | 12 |
| 1 | 李祎, 林振泉, 刘子鹤. 酿酒酵母适应性实验室进化工具的最新进展[J]. 合成生物学, 2021, 2(2): 287-301. |
| LI Y, LIN Z Q, LIU Z H. Advances in yeast based adaptive laboratory evolution[J]. Synthetic Biology Journal, 2021, 2(2): 287-301. | |
| 2 | 夏思杨, 江丽红, 蔡谨, 等. 酿酒酵母基因组进化的研究进展[J]. 合成生物学, 2020, 1(5): 556-569. |
| XIA S Y, JIANG L H, CAI J, et al. Advances in genome evolution of Saccharomyces cerevisiae [J]. Synthetic Biology Journal, 2020, 1(5): 556-569. | |
| 3 | SI T, XIAO H, ZHAO H M. Rapid prototyping of microbial cell factories via genome-scale engineering[J]. Biotechnology Advances, 2015, 33(7): 1420-1432. |
| 4 | LUAN G D, CAI Z, LI Y, et al. Genome replication engineering assisted continuous evolution (GREACE) to improve microbial tolerance for biofuels production[J]. Biotechnology for Biofuels, 2013, 6(1): 137. |
| 5 | BADRAN A H, LIU D R. In vivo continuous directed evolution[J]. Current Opinion in Chemical Biology, 2015, 24: 1-10. |
| 6 | ARNOLD F H, VOLKOV A A. Directed evolution of biocatalysts[J]. Current Opinion in Chemical Biology, 1999, 3(1): 54-59. |
| 7 | WANG Y J, XUE P, CAO M F, et al. Directed evolution: methodologies and applications[J]. Chemical Reviews, 2021, 121(20): 12384-12444. |
| 8 | 祁延萍, 朱晋, 张凯, 等. 定向进化在蛋白质工程中的应用研究进展 [J]. 合成生物学, 2022, 3(6): 1-29. |
| QI Y P, ZHU J, ZHANG K, et al. Recent development of directed evolution in protein engineering [J]. Synthetic Biology Journal, 2022, 3(6): 1-29. | |
| 9 | SCHELLER J, SCHÜRER A, RUDOLPH C, et al. MPH1, a yeast gene encoding a DEAH protein, plays a role in protection of the genome from spontaneous and chemically induced damage[J]. Genetics, 2000, 155(3): 1069-1081. |
| 10 | SPEYER J F. Mutagenic DNA polymerase[J]. Biochemical and Biophysical Research Communications, 1965, 21(1): 6-8. |
| 11 | CHOU H H, KEASLING J D. Programming adaptive control to evolve increased metabolite production[J]. Nature Communications, 2013, 4: 2595. |
| 12 | DROTSCHMANN K, CLARK A B, TRAN H T, et al. Mutator phenotypes of yeast strains heterozygous for mutations in the MSH2 gene[J]. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(6): 2970-2975. |
| 13 | SERERO A, JUBIN C, LOEILLET S, et al. Mutational landscape of yeast mutator strains[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(5): 1897-1902. |
| 14 | REENAN R A, KOLODNER R D. Characterization of insertion mutations in the Saccharomyces cerevisiae MSH1 and MSH2 genes: evidence for separate mitochondrial and nuclear functions[J]. Genetics, 1992, 132(4): 975-985. |
| 15 | KATJU V, KONRAD A, DEISS T C, et al. Mutation rate and spectrum in obligately outcrossing Caenorhabditis elegans mutation accumulation lines subjected to RNAi-induced knockdown of the mismatch repair gene msh-2 [J]. G3 Genes|Genomes|Genetics, 2021, 12(1): jkab364. |
| 16 | GOTO M, SASAKI M, KOBAYASHI T. The S-phase cyclin Clb5 promotes rRNA gene (rDNA) stability by maintaining replication initiation efficiency in rDNA[J]. Molecular and Cellular Biology, 2021, 41(5): e00324-e00320. |
| 17 | TISHKOFF D X, FILOSI N, GAIDA G M, et al. A novel mutation avoidance mechanism dependent on S. cerevisiae RAD27 is distinct from DNA mismatch repair[J]. Cell, 1997, 88(2): 253-263. |
| 18 | MCCULLEY J L, PETES T D. Chromosome rearrangements and aneuploidy in yeast strains lacking both Tel1p and Mec1p reflect deficiencies in two different mechanisms[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(25): 11465-11470. |
| 19 | PENNANEACH V, KOLODNER R D. Recombination and the Tel1 and Mec1 checkpoints differentially effect genome rearrangements driven by telomere dysfunction in yeast[J]. Nature Genetics, 2004, 36(6): 612-617. |
| 20 | PARK H, KIM S. Gene-specific mutagenesis enables rapid continuous evolution of enzymes in vivo [J]. Nucleic Acids Research, 2021, 49(6): e32. |
| 21 | YONA A H, MANOR Y S, HERBST R H, et al. Chromosomal duplication is a transient evolutionary solution to stress[J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(51): 21010-21015. |
| 22 | GILBERT L A, LARSON M H, MORSUT L, et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes[J]. Cell, 2013, 154(2): 442-451. |
| 23 | YAO Z, WANG Q H, DAI Z J. Recent advances in directed yeast genome evolution[J]. Journal of Fungi, 2022, 8(6): 635. |
| 24 | LIAN J Z, HAMEDIRAD M, HU S M, et al. Combinatorial metabolic engineering using an orthogonal tri-functional CRISPR system[J]. Nature Communications, 2017, 8: 1688. |
| 25 | GIETZ R D, SCHIESTL R H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method[J]. Nature Protocols, 2007, 2(1): 31-34. |
| 26 | PATTERSON M N, MAXWELL P H. Combining magnetic sorting of mother cells and fluctuation tests to analyze genome instability during mitotic cell aging in Saccharomyces cerevisiae [J]. Journal of Visualized Experiments: JoVE, 2014(92): e51850. |
| 27 | BOEKE J D, LACROUTE F, FINK G R. A positive selection for mutants lacking orotidine-5'-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance[J]. Molecular & General Genetics: MGG, 1984, 197(2): 345-346. |
| 28 | WHELAN W L, GOCKE E, MANNEY T R. The CAN1 locus of Saccharomyces cerevisiae: fine-structure analysis and forward mutation rates[J]. Genetics, 1979, 91(1): 35-51. |
| 29 | QI M M, ZHANG B, JIANG L H, et al. PCR & Go: a pre-installed expression chassis for facile integration of multi-gene biosynthetic pathways[J]. Frontiers in Bioengineering and Biotechnology, 2021, 8: 613771. |
| 30 | TSAI C S, KONG I I, LESMANA A, et al. Rapid and marker-free refactoring of xylose-fermenting yeast strains with Cas9/CRISPR[J]. Biotechnology and Bioengineering, 2015, 112(11): 2406-2411. |
| 31 | 丁明珠, 李炳志, 王颖, 等. 合成生物学重要研究方向进展[J]. 合成生物学, 2020, 1(1): 7-28. |
| DING M Z, LI B Z, WANG Y, et al. Significant research progress in synthetic biology[J]. Synthetic Biology Journal, 2020, 1(1): 7-28. |
| [1] | 孙文涛, 张昕哲, 万盛通, 王茹雯, 李春. Ⅱ型细胞色素P450酶氧化β-香树脂醇的选择性调控研究[J]. 合成生物学, 2021, 2(5): 804-814. |
| [2] | 李晓东, 杨成帅, 王平平, 严兴, 周志华. 构建酿酒酵母细胞工厂从头合成倍半萜类化合物α-新丁香三环烯和β-石竹烯[J]. 合成生物学, 2021, 2(5): 792-803. |
| [3] | 李祎, 林振泉, 刘子鹤. 酿酒酵母适应性实验室进化工具的最新进展[J]. 合成生物学, 2021, 2(2): 287-301. |
| [4] | 盛月, 张根林. 酵母终止子工程:从机理探索到人工设计[J]. 合成生物学, 2020, 1(6): 709-721. |
| [5] | 夏思杨, 江丽红, 蔡谨, 黄磊, 徐志南, 连佳长. 酿酒酵母基因组进化的研究进展[J]. 合成生物学, 2020, 1(5): 556-569. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
