|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
合成基因线路的工程化设计研究进展与展望
合成生物学
2025, 6 (1):
45-64.
DOI:10.12211/2096-8280.2023-096
合成基因线路利用合成生物学的技术和方法,将生物元件进行重新设计与构建,使人工设计的生物分子线路在活细胞中行使特定生物功能,在生物制造、医疗健康以及环境监测等领域具有巨大的潜力。但其工程化设计仍受到各种因素的制约,包括正交元器件数量有限、大规模线路组装困难、线路行为预测性低等。根据研究者们开发的各种调控元件工具箱和组装方法,本文逐点阐述了工程化设计基因线路所需遵循的几个核心原则:正交化、标准化、模块化与自动化。文章从DNA复制、转录和翻译层面介绍了正交基因元件库的构建和改造方法;全面总结了基因元件的标准化定量表征方法与标准元件设计方法;并介绍了本团队与其他团队在模块化基因线路设计方面的相关进展;分别从软件、硬件和人工智能角度展示如何实现基因线路的自动化设计。最后,本文探讨了基因线路设计的未来发展趋势,指出需要进一步融合人工智能和自动化等信息技术来加速基因线路“设计-构建-测试-学习”循环的迭代,提高线路设计的功能可预测性和复杂性,高效设计出符合目标需求的人造生命体。
表1
典型正交基因元件库的设计与表征
正文中引用本图/表的段落
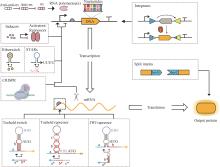
生物正交反应是指能够在生物体系中进行,且不会与天然生物化学过程相互干扰的一类化学反应[13]。在基因线路设计过程中,为了精准地调控细胞内各组分的活性,应尽量减少元件之间的相互干扰,这就是基因线路中的“正交化”。随着合成生物学的快速发展,科学家们在遗传信息表达及生化代谢的不同过程中成功创建了正交元件(图1和表1)。
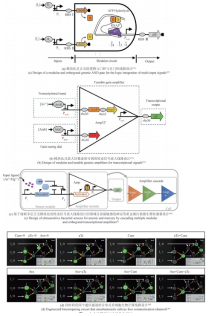
同时,单个细胞的负载有限,组合多种功能细胞,可以提高群体对复杂环境的适应性和鲁棒性,以完成更加复杂的工作。可通过多细胞分布式线路设计,让单个细胞里的线路模块化并进行多个细胞间的级联通信来提高线路的模块性和实现线路的规模化组装设计。相比于基于单种细胞的计算线路,分布式计算不仅可以降低每个细胞中合成线路的体积,减少代谢负担,而且可以利用细胞高度模块化的特性,在不同的细胞群体内重复使用相同的基因元件,降低对正交基因调控元件的依赖。为了演示更复杂的多细胞生物计算功能,研究人员设计了复杂的三输入XOR-AND逻辑门电路。XOR-AND逻辑门电路分别部署在七个不同的大肠杆菌菌株中,由四个通信通道协调。每个菌株包含一个NOR门(cell-1至cell-6)或一个Buffer门(cell-7),这是已知第一个同时利用四个通信通道的生物计算线路[图3(d)][38]。虽然多细胞分布式计算在细菌、酵母和哺乳动物细胞中都已经实现,但它们的规模却并未得到显著的提高。在细菌中最具有代表性的分布式计算线路仍然是基于多细胞或非门的16个2输入逻辑门以及更复杂一些的与门-异或门复合逻辑[3, 38]。而在酵母与哺乳动物细胞中,分布式计算线路的规模也与单细胞计算线路的规模相差不大,最具代表性的是1位全加器程序[90-91]。
本文的其它图/表
|