|
||
|
合成基因线路的工程化设计研究进展与展望
合成生物学
2025, 6 (1):
45-64.
DOI:10.12211/2096-8280.2023-096
合成基因线路利用合成生物学的技术和方法,将生物元件进行重新设计与构建,使人工设计的生物分子线路在活细胞中行使特定生物功能,在生物制造、医疗健康以及环境监测等领域具有巨大的潜力。但其工程化设计仍受到各种因素的制约,包括正交元器件数量有限、大规模线路组装困难、线路行为预测性低等。根据研究者们开发的各种调控元件工具箱和组装方法,本文逐点阐述了工程化设计基因线路所需遵循的几个核心原则:正交化、标准化、模块化与自动化。文章从DNA复制、转录和翻译层面介绍了正交基因元件库的构建和改造方法;全面总结了基因元件的标准化定量表征方法与标准元件设计方法;并介绍了本团队与其他团队在模块化基因线路设计方面的相关进展;分别从软件、硬件和人工智能角度展示如何实现基因线路的自动化设计。最后,本文探讨了基因线路设计的未来发展趋势,指出需要进一步融合人工智能和自动化等信息技术来加速基因线路“设计-构建-测试-学习”循环的迭代,提高线路设计的功能可预测性和复杂性,高效设计出符合目标需求的人造生命体。 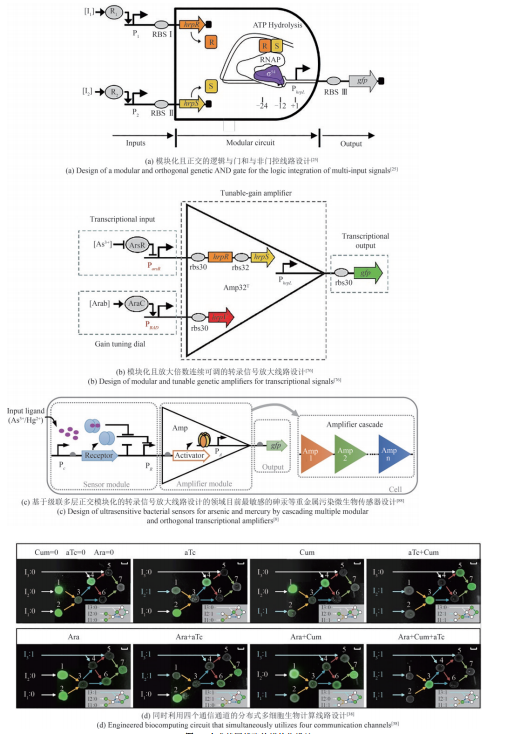 View image in article
图3
合成基因线路的模块化设计
正文中引用本图/表的段落
模块化、不受细胞类型影响且可编程的系统是合成生物学研究中的重要瓶颈问题,相关的研究不断在扩充模块化的基因线路,使其不受转录、翻译资源影响,具备可预测性。本团队在标准化工作环境下充分测试基因线路各组成部件特性,利用铜绿假单胞菌中的异源σ54依赖型转录因子和相关基因调控元件在大肠杆菌中率先设计并构建了模块化且正交化的逻辑与门和与非门控线路、模块化且放大倍数连续可调的转录信号放大线路[图3(a)和(b)][25, 76]。并通过结合现有的sgRNA支架设计和一种已被证明模块性较高的σ54因子依赖型激活子PspF,在模式生物大肠杆菌(Escherichia coli)和固氮菌(Klebsiella oxytoca)中实现了类真核CRISPR激活,并且激活装置具有泄露表达水平低、激活倍率高等优异的调控性能[45]。经过进一步的工程化改造和优化,其基因调控性能第一次达到了足以构建CRISPRa级联调控、CRISPRa正反馈调控和多通道基因调控的水平上。Li等[77]在哺乳动物细胞中结合新设计的杂交启动子并通过关键转录起始元件的空间位阻发挥转录抑制作用,构建了包含26种可逆转录激活因子样效应抑制因子(transcription activator-like effector repressors,TALER)的文库,可以准确预测模块化组装的TALER级联和开关电路的行为。很多天然的转录因子已经被深入研究并应用于代谢通路动态调控和人工基因线路构建[78]。然而,天然的转录因子的调控性能难以满足工程化设计的需求,传统筛选和定向进化方法可改善转录因子的性能,但相关实验周期长、工作量大。Hou等[79]针对转录因子的协同性、超敏性需求,开发了一种模块化的协同性蛋白元件,将该元件与转录因子融合后,可有效提高转录因子结合DNA的协同性,从而显著提升转录调控系统的超敏性和严谨性。该元件普遍适用于各种转录因子,在不同物种中也具有很高的通用性,可对遗传开关和人工调控网络实行“即插即用”式的改造,大幅改善人工系统的性能。
复杂基因线路中基因模块的行为通常是不可预测的[80]。在组装模块为基因线路后,下游元件(例如调节蛋白的结合位点)会对上游模块产生负载,从而对预测的行为产生干扰[81-83]。Ceroni等[84]开发了一套用单位细胞产生的荧光量反映人工基因元件对细胞资源的占用情况的检测方法,该方法首次建立模型来计算可使用的细胞资源,有助于选择出最优的基因线路设计方案。细胞生长的内部环境是动态变化的,这使得构建基因线路或者网络时,在细胞中引入的元件难以实现稳定的输出,如众所熟知的启动子。MIT Voigt实验室的研究人员运用控制论理论设计了能够在任何拷贝数下实现目的基因同一水平表达的启动子元件,研究人员利用transcription-activator-like effector(TALE)构建了iFFL稳定的启动子(iFFLs-stabilized promoter)。结果表明,即使基因的拷贝数由于基因组突变或者培养基条件改变发生了变化,这些iFFL稳定的启动子控制的目的基因在不同的基因组位点或者质粒上仍能实现接近相同的表达水平[85]。本团队首次证明了人工合成核酸海绵可系统地调节基因线路中的基因表达,从而精确改变该线路中的基因表达泄露、输出幅度和诱导倍数、对小分子的响应灵敏度,并改善宿主细胞的生长速度[86]。该基因调节方法简单、有效,可广泛用于多种应用领域相关的人工合成基因线路设计。Mishra等[87]设计了一种包含快速磷酸转移过程的负载驱动器,即使在存在大量负载的情况下也能响应时变输入信号,并且在酿酒酵母中测试了该模块的有效性。该负载驱动器可作为创建复杂的、更高级别的基因线路的基本构建块。本团队通过级联多层正交模块化的转录信号放大线路,首先通过调节细胞内受体蛋白的密度来提高传感器的灵敏度,其次通过串联多极转录放大器来促进输出模块的信号强度,最后结合转录调节和翻译后调节的方法来降低在无输入信号时传感器的信号输出,从而增加输出信号的动态范围,创建了领域目前最敏感的基于大肠杆菌的砷和汞等重金属污染细胞传感器及创新的可视化输出模块[图3(c)][88]。Liu等[89]揭示了Ⅱ型CRISPR系统crRNA-tracrRNA配对的可编程性机制,利用重编程的tracrRNA劫持各种RNA分子激活CRISPR系统,并利用双gRNA介导的CRISPR激活(CRISPRa)系统,构建了一系列正交化的可编程AND-gate布尔逻辑门库,为编程生物学带来了更多可能性,包括重组细胞内的基因调控网络拓扑结构、开发新的RNA编辑器和RNA标记方法、设计复杂细胞生物计算线路等。
同时,单个细胞的负载有限,组合多种功能细胞,可以提高群体对复杂环境的适应性和鲁棒性,以完成更加复杂的工作。可通过多细胞分布式线路设计,让单个细胞里的线路模块化并进行多个细胞间的级联通信来提高线路的模块性和实现线路的规模化组装设计。相比于基于单种细胞的计算线路,分布式计算不仅可以降低每个细胞中合成线路的体积,减少代谢负担,而且可以利用细胞高度模块化的特性,在不同的细胞群体内重复使用相同的基因元件,降低对正交基因调控元件的依赖。为了演示更复杂的多细胞生物计算功能,研究人员设计了复杂的三输入XOR-AND逻辑门电路。XOR-AND逻辑门电路分别部署在七个不同的大肠杆菌菌株中,由四个通信通道协调。每个菌株包含一个NOR门(cell-1至cell-6)或一个Buffer门(cell-7),这是已知第一个同时利用四个通信通道的生物计算线路[图3(d)][38]。虽然多细胞分布式计算在细菌、酵母和哺乳动物细胞中都已经实现,但它们的规模却并未得到显著的提高。在细菌中最具有代表性的分布式计算线路仍然是基于多细胞或非门的16个2输入逻辑门以及更复杂一些的与门-异或门复合逻辑[3, 38]。而在酵母与哺乳动物细胞中,分布式计算线路的规模也与单细胞计算线路的规模相差不大,最具代表性的是1位全加器程序[90-91]。
本文的其它图/表
|


{kind=link}