| 微生物种类 | 常见产油菌种 | 油脂含量 (占干重百分比) | 主要脂肪酸 组成 | 特征 | 参考 文献 |
|---|
| 酵母菌 | 解脂耶氏酵母 Y. lipolytica | 通常在20%-45%之间,特定条件下可超过90% | 油酸、棕榈酸、棕榈油酸 | 遗传改造技术成熟;底物利用谱广,可代谢乙酸、脂肪酸、油脂、正烷烃;抗逆性较强 | [12-15] | | 圆红冬孢酵母 R. toruloides | 通常在40%-60%之间,可超过70% | 油酸、棕榈酸、硬脂酸 | 遗传工具相对丰富,可利用农林废弃物产油 | [16-19] | | 产油丝孢酵母 C. oleaginosus | 通常大于50% | 油酸、棕榈酸、硬脂酸 | 代谢木糖能力强;能利用木质纤维素水解液及工业废弃物 | [16-18, 20, 21] | | 斯达氏油脂酵母 L. starkeyi | 通常为30%-60%, 可超过70% | 油酸、棕榈酸、硬脂酸 | 能够代谢木糖;遗传工具较少 | [16, 17, 20] | | 霉菌 | 高山被孢霉 M. alpina | 通常在40%-60%之间,可超过70% | 油酸、棕榈酸、ARA | 商业化生产ARA的主要菌株 | [22-24] | | 卷枝毛霉 M. circinelloides | 通常为25%-53% | 亚油酸、油酸、GLA | GLA含量高;调控机制研究较多;遗传改造技术相对丰富;代谢工程潜力大;可利用葡萄糖、乙酸等做碳源 | [25, 26] | | 藻类 | 裂殖壶菌 A. limacinum | 通常在30%-50%之间,可超过60% | DHA、棕榈酸 | 生长速度快、发酵密度高;高产DHA;油脂类型适合婴幼儿补充 | [27, 28] | | 小球藻 C.vulgaris | 通常为41%-58% | 棕榈酸、油酸、亚油酸、ALA、GLA | ALA和GLA含量高,适宜用于食品补充剂;能够用来处理城市及工业废水 | [29, 30] | | 三角褐指藻 P. tricornutum | 通常为18%-57% | EPA、棕榈酸、棕榈油酸 | EPA产量高,不饱和脂肪酸含量尤其是ω3/ω6比值高,油脂营养价值较好 | [29, 31] | | 细菌 | 浑浊红球菌 R. opacus | 通常在20%-40%之间,可超过50% | 棕榈酸、 十七烷酸、油酸 | 超长链脂肪酸产量高;可利用木质纤维素水解液及工业废弃物 | [11, 32, 33] | | 甲基微菌 M. buryatense | 通常为10%-30% | 中短链脂肪酸 | 可以甲烷为碳源合成脂肪酸,生物转化一碳化合物潜力大 | [34] |
Table 1
Summary of oil-producing microbial resources and their characteristics
Extracts from the Article
产油微生物通常指能够累积超过细胞干重20%油脂的微生物,具有独特的代谢特征,在高碳氮比条件下,乙酰辅酶A和NADPH向脂肪酸和甘油三酯合成途径通量[10, 11]。产油微生物涉及酵母、霉菌、微藻和细菌等类群(表1)。
大豆是一种豆科蔬菜作物,近年来,我国对豆油和豆粕的需求量迅速增加。此外,大豆还是重要的蛋白质来源,在食品和制药工业有重要应用[55]。根据美国农业部的数据,截止2024年10月,世界大豆油的产量约为6.6亿吨。大豆油含量约占大豆种子重量的17%~23%,这相较其他油料作物的比例偏低,如花生种子的含油量约为45%。目前在提升大豆总含油量方面的进展有限,主要原因是需要做到在提升油脂含量的同时,不影响其蛋白含量[56]。因此,受两用价值的影响,在不进一步扩大种植面积的情况下,大豆油产量难以得到显著提升。但随着研究的不断深入,尤其是分子技术、各类组学技术的不断发展,在提高大豆含油量的研究中也取得了不错的进展。据报道,将拉曼伞形霉来源的二酰甘油酰基转移酶2A (DGAT2A) 在大豆中表达,可使大豆种子的含油量提升1.5%,重要的是,得到的转基因大豆的产量和蛋白质含量均未受到显著影响[57]。目前,已经在大豆种子中鉴定出多个油脂合成相关的重要转录因子,如GmbZIP123可调节蔗糖转运体和细胞壁转化酶的表达来调控糖转运;GmDofs可通过激活乙酰辅酶A羧化酶(ACCase)或长链脂酰辅酶A合成酶(LACS)等途径参与脂肪酸合成调控;GmMYB73能够抑制油分累积负调控因子GL2(Glabra 2)的表达,从而解除对磷脂酶PLDα1的抑制,促进甘油二酯和甘油三酯的合成[58-61]。
基于对各项指标的综合分析与比较,植物油脂的生产成本较低,这主要得益于成熟的种植和加工技术.根据马来西亚棕榈油局(Malaysian Palm Oil Board, MPOB)的报告测算,棕榈油的生产成本可低于4000元/吨,是目前生产成本最低的植物油脂[ 52];菜籽油的生产成本最高,根据2021年加拿大油菜籽市场报告《Canola Oil Production Costs》统计数据预测,菜籽油的生产成本不小于7000元/吨;大豆油和葵花籽油的生产成本介于两者之间.微生物油脂的生产依赖发酵工艺和碳源,目前成本较高,由于尚缺乏规模化生产,难以严格核算成本,但依据中试规模的核算,行业内通常认为目前微生物油脂的生产成本高于20000元/吨.根据行业调研分析,近年来,裂殖壶菌产业化生产DHA的单位成本随着技术进步和工艺优化有所下降,2020年的单位成本约为520000元/吨.异养微藻发酵的培养条件较常规微生物更为复杂,需要碳源、氮源、pH值、温度、溶氧等条件的精准控制及严格的无菌条件,这无疑增加了一定的生产成本[ 53]. ... Kadir APG, Oil palm economic performance in Malaysia and R&D progress in 2019 1 ... 基于对各项指标的综合分析与比较,植物油脂的生产成本较低,这主要得益于成熟的种植和加工技术.根据马来西亚棕榈油局(Malaysian Palm Oil Board, MPOB)的报告测算,棕榈油的生产成本可低于4000元/吨,是目前生产成本最低的植物油脂[52];菜籽油的生产成本最高,根据2021年加拿大油菜籽市场报告《Canola Oil Production Costs》统计数据预测,菜籽油的生产成本不小于7000元/吨;大豆油和葵花籽油的生产成本介于两者之间.微生物油脂的生产依赖发酵工艺和碳源,目前成本较高,由于尚缺乏规模化生产,难以严格核算成本,但依据中试规模的核算,行业内通常认为目前微生物油脂的生产成本高于20000元/吨.根据行业调研分析,近年来,裂殖壶菌产业化生产DHA的单位成本随着技术进步和工艺优化有所下降,2020年的单位成本约为520000元/吨.异养微藻发酵的培养条件较常规微生物更为复杂,需要碳源、氮源、pH值、温度、溶氧等条件的精准控制及严格的无菌条件,这无疑增加了一定的生产成本[53]. ... Towards maximizing biomass and lipid productivity: high-throughput screening assay for prospecting heterotrophic growth for new microalgal isolates 1 2024 ... 基于对各项指标的综合分析与比较,植物油脂的生产成本较低,这主要得益于成熟的种植和加工技术.根据马来西亚棕榈油局(Malaysian Palm Oil Board, MPOB)的报告测算,棕榈油的生产成本可低于4000元/吨,是目前生产成本最低的植物油脂[52];菜籽油的生产成本最高,根据2021年加拿大油菜籽市场报告《Canola Oil Production Costs》统计数据预测,菜籽油的生产成本不小于7000元/吨;大豆油和葵花籽油的生产成本介于两者之间.微生物油脂的生产依赖发酵工艺和碳源,目前成本较高,由于尚缺乏规模化生产,难以严格核算成本,但依据中试规模的核算,行业内通常认为目前微生物油脂的生产成本高于20000元/吨.根据行业调研分析,近年来,裂殖壶菌产业化生产DHA的单位成本随着技术进步和工艺优化有所下降,2020年的单位成本约为520000元/吨.异养微藻发酵的培养条件较常规微生物更为复杂,需要碳源、氮源、pH值、温度、溶氧等条件的精准控制及严格的无菌条件,这无疑增加了一定的生产成本[53]. ... Exploring the future of vegetable oils : oil crop implications : fats 1 2024 ... 大宗油脂是指在生产规模、消费量和市场影响力等方面占据主导地位的油脂,油料作物是大宗油脂的主要来源.世界自然保护联盟(International Union for Conservation of Nature, IUCN)最新报告显示,截止2024年,全球油料作物占地约5.4亿公顷,约为农业总面积的37%[54].近年来,随着生物技术的发展,尤其是基因工程技术的发展,结合常规的育种手段,为油料作物提供了高质量的种子,在一定程度上也提高了大宗油脂的产量.然而,对于油脂的需求仍在进一步增加,IUCN的报告指出,目前有28%的油料作物被用于生物柴油、动物饲料和工业原料.即使将产出的植物油完全用于食品工业,到2050年全球植物油产量仍需提高14%左右才能满足约97亿人口的需求.目前大豆油、棕榈油和菜籽油占据全球植物油市场的主导地位,根据联合国粮食及农业组织(Food and Agriculture Organization of the United Nations, FAO)2022年的统计数据,其总产量超过全球油脂的75%,其中美国、巴西和阿根廷是主要的大豆油生产国;印度尼西亚和马来西亚是全球最大的棕榈油生产国;中国、加拿大和欧盟国家主导菜籽油生产. ... Massive soybean expansion in South America since 2000 and implications for conservation 1 2021 ... 大豆是一种豆科蔬菜作物,近年来,我国对豆油和豆粕的需求量迅速增加.此外,大豆还是重要的蛋白质来源,在食品和制药工业有重要应用[55].根据美国农业部的数据,截止2024年10月,世界大豆油的产量约为6.6亿吨.大豆油含量约占大豆种子重量的17%~23%,这相较其他油料作物的比例偏低,如花生种子的含油量约为45%.目前在提升大豆总含油量方面的进展有限,主要原因是需要做到在提升油脂含量的同时,不影响其蛋白含量[56].因此,受两用价值的影响,在不进一步扩大种植面积的情况下,大豆油产量难以得到显著提升.但随着研究的不断深入,尤其是分子技术、各类组学技术的不断发展,在提高大豆含油量的研究中也取得了不错的进展.据报道,将拉曼伞形霉来源的二酰甘油酰基转移酶2A (DGAT2A) 在大豆中表达,可使大豆种子的含油量提升1.5%,重要的是,得到的转基因大豆的产量和蛋白质含量均未受到显著影响[57].目前,已经在大豆种子中鉴定出多个油脂合成相关的重要转录因子,如GmbZIP123可调节蔗糖转运体和细胞壁转化酶的表达来调控糖转运;GmDofs可通过激活乙酰辅酶A羧化酶(ACCase)或长链脂酰辅酶A合成酶(LACS)等途径参与脂肪酸合成调控;GmMYB73能够抑制油分累积负调控因子GL2(Glabra 2)的表达,从而解除对磷脂酶PLDα1的抑制,促进甘油二酯和甘油三酯的合成[58-61]. ... genetic approaches for modification of functionality and total content 1 2009 ... 大豆是一种豆科蔬菜作物,近年来,我国对豆油和豆粕的需求量迅速增加.此外,大豆还是重要的蛋白质来源,在食品和制药工业有重要应用[55].根据美国农业部的数据,截止2024年10月,世界大豆油的产量约为6.6亿吨.大豆油含量约占大豆种子重量的17%~23%,这相较其他油料作物的比例偏低,如花生种子的含油量约为45%.目前在提升大豆总含油量方面的进展有限,主要原因是需要做到在提升油脂含量的同时,不影响其蛋白含量[56].因此,受两用价值的影响,在不进一步扩大种植面积的情况下,大豆油产量难以得到显著提升.但随着研究的不断深入,尤其是分子技术、各类组学技术的不断发展,在提高大豆含油量的研究中也取得了不错的进展.据报道,将拉曼伞形霉来源的二酰甘油酰基转移酶2A (DGAT2A) 在大豆中表达,可使大豆种子的含油量提升1.5%,重要的是,得到的转基因大豆的产量和蛋白质含量均未受到显著影响[57].目前,已经在大豆种子中鉴定出多个油脂合成相关的重要转录因子,如GmbZIP123可调节蔗糖转运体和细胞壁转化酶的表达来调控糖转运;GmDofs可通过激活乙酰辅酶A羧化酶(ACCase)或长链脂酰辅酶A合成酶(LACS)等途径参与脂肪酸合成调控;GmMYB73能够抑制油分累积负调控因子GL2(Glabra 2)的表达,从而解除对磷脂酶PLDα1的抑制,促进甘油二酯和甘油三酯的合成[58-61]. ... Expression of Umbelopsis ramanniana DGAT2A in seed increases oil in soybean 2 2008 ... 大豆是一种豆科蔬菜作物,近年来,我国对豆油和豆粕的需求量迅速增加.此外,大豆还是重要的蛋白质来源,在食品和制药工业有重要应用[55].根据美国农业部的数据,截止2024年10月,世界大豆油的产量约为6.6亿吨.大豆油含量约占大豆种子重量的17%~23%,这相较其他油料作物的比例偏低,如花生种子的含油量约为45%.目前在提升大豆总含油量方面的进展有限,主要原因是需要做到在提升油脂含量的同时,不影响其蛋白含量[56].因此,受两用价值的影响,在不进一步扩大种植面积的情况下,大豆油产量难以得到显著提升.但随着研究的不断深入,尤其是分子技术、各类组学技术的不断发展,在提高大豆含油量的研究中也取得了不错的进展.据报道,将拉曼伞形霉来源的二酰甘油酰基转移酶2A (DGAT2A) 在大豆中表达,可使大豆种子的含油量提升1.5%,重要的是,得到的转基因大豆的产量和蛋白质含量均未受到显著影响[57].目前,已经在大豆种子中鉴定出多个油脂合成相关的重要转录因子,如GmbZIP123可调节蔗糖转运体和细胞壁转化酶的表达来调控糖转运;GmDofs可通过激活乙酰辅酶A羧化酶(ACCase)或长链脂酰辅酶A合成酶(LACS)等途径参与脂肪酸合成调控;GmMYB73能够抑制油分累积负调控因子GL2(Glabra 2)的表达,从而解除对磷脂酶PLDα1的抑制,促进甘油二酯和甘油三酯的合成[58-61]. ...
Other Images/Table from this Article
-
 Table 2
Analysis of economic difference between vegetable oil and microbial oil
Table 2
Analysis of economic difference between vegetable oil and microbial oil
-
 Fig. 1
Metabolic pathways for microbial synthesis of various long-chain/very long-chain unsaturated fatty acids(OA, oleic acid; LA, linoleic acid; ALA, alpha-linolenic acid; GLA, gamma-linolenic acid; EDA, eicosadienoic acid; DGLA, dihomo-gamma-linolenic acid; STA, stearidonic acid; ETrA, eicosatrienoic acid; ETA, eicosatetraenoic acid; DPA, docosapentaenoic acid; Des, desaturase; Elo, elongase; KCS, 3-ketoacyl-CoA synthase. The dashed line represents a multi-step reaction)
Fig. 1
Metabolic pathways for microbial synthesis of various long-chain/very long-chain unsaturated fatty acids(OA, oleic acid; LA, linoleic acid; ALA, alpha-linolenic acid; GLA, gamma-linolenic acid; EDA, eicosadienoic acid; DGLA, dihomo-gamma-linolenic acid; STA, stearidonic acid; ETrA, eicosatrienoic acid; ETA, eicosatetraenoic acid; DPA, docosapentaenoic acid; Des, desaturase; Elo, elongase; KCS, 3-ketoacyl-CoA synthase. The dashed line represents a multi-step reaction)
-
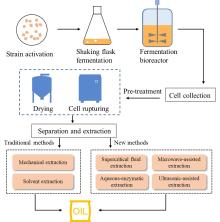 Fig. 2
Production process of microbial oil
Fig. 2
Production process of microbial oil
|