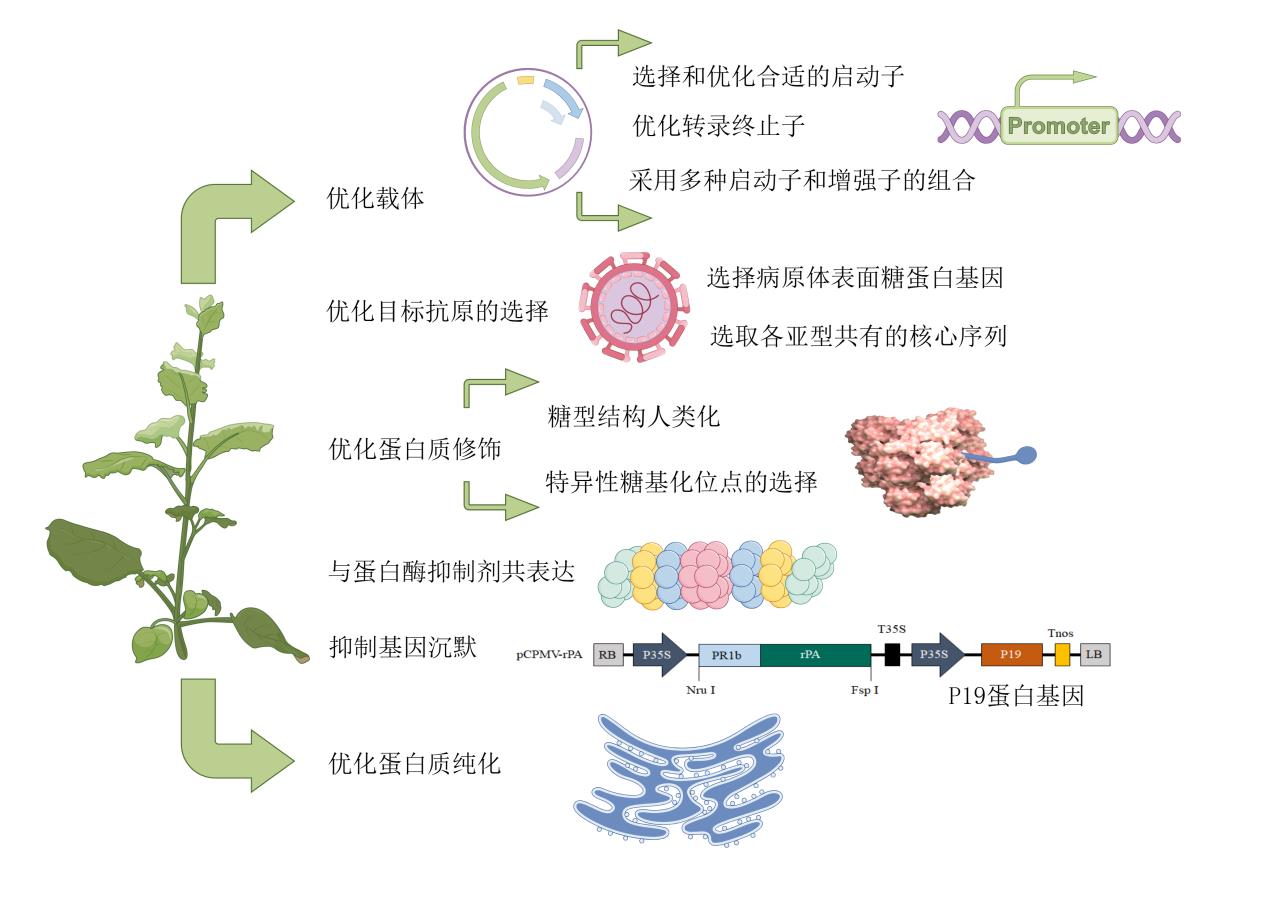
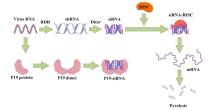
基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152]。马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153]。在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示。Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155]。此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156]。而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制。其具体抑制机制还有待深入研究。小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能。在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]。
沉默抑制技术主要针对植物细胞内的基因沉默机制,但其对疫苗抗原表达的直接影响尚不完全明确.在某些情况下,沉默抑制可能会对疫苗抗原的表达产生负面影响,如改变抗原的糖基化修饰模式、影响抗原的折叠和组装等,从而影响疫苗的免疫原性和保护效果. ...
Application of Baculovirus Expression Vector System (BEVS) in Vaccine Development
1
2023
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
聚合酶Ⅱ转录的sRNA与RNA沉默抑制子对TMV病毒载体表达系统作用的比较
1
2016
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
Comparison of the Effects of Pol II-transcribed sRNA and RNA Silencing Suppressors on the TMV Viral Vector Expression System
1
2016
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
The Tombusvirus-encoded P19: from irrelevance to elegance
1
2006
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
Co-expression With Replicating Vector Overcoming Competitive Effects Derived by a Companion Protease Inhibitor in Plants
1
2021
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
芸薹黄化病毒P0蛋白抑制RNA沉默的分子机制研究
1
2015
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
Molecular Mechanism of RNA Silencing Suppression by P0 Protein of Brassica Yellows Virus
1
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
小麦黄花叶病毒P1蛋白具有RNA沉默抑制活性促进病毒侵染小麦
1
2023
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
The P1 protein of wheat yellow mosaic virus exerts RNA silencing suppression activity to facilitate virus infection in wheat
1
2023
... 基因沉默抑制是植物源疫苗研究中的关键环节,它可以帮助调控目标基因的表达,从而优化疫苗的产量和质量[117, 152].马婷等在2016年体外合成了拟南芥U6-1 RNA基因和黄瓜花叶病毒(CMV)编码的沉默抑制蛋白2b基因,并通过农杆菌浸润法接种烟草,推测出GFP基因表达增强是由于核外输出的竞争导致输出到细胞质的保护性抗病毒mRNA减少,从而为改进植物源重组蛋白表达提供了新的思路[153].在植物源疫苗中,还存在一种高效的基因沉默抑制机制,即番茄丛矮病毒编码的P19蛋白,通过与siRNA结合从而抑制RISC复合体的形成,致使其无法特异性切割与该siRNA同源的靶mRNA[154],如图2所示.Jiexue等人的研究中采用过P19蛋白共表达目的基因 [155].此外,马铃薯卷叶病毒属编码的一种沉默抑制蛋白P0可以通过SKP1-CULLIN1复合体将一种RISC复合体——AGO1蛋白泛素化,从而介导AGO1的降解,AGO1的与核酸的结合活性也会受到其它沉默蛋白的影响,如AGO1的PAZ区与CMV编码的2b蛋白的结合会抑制AGO1与核酸链的结合[156].而AGO1的降解或结合活性的减弱会导致基因沉默的过程受到抑制.其具体抑制机制还有待深入研究.小麦黄花叶病毒(WYMV)编码的P1蛋白和P38蛋白机制类似,通过与AGO1互作来行使沉默抑制功能.在陈道等人的实验中,P1蛋白通过干扰钙调蛋白相关的抗病毒 RNAi 防御来充当 VSR,进而促进病毒侵染[157]. ...
植物表达外源蛋白研究进展及展望
1
2023
... 外源蛋白在植物细胞内还可能引起内质网胁迫,影响蛋白的正确折叠和积累,该情况可以通过优化内质网蛋白折叠与质量监控系统和增强内质网相关降解ERAD 途径来处理和清除内质网中错误折叠的有缺陷的蛋白质,可以提高外源蛋白的表达效率[158],维持内质网稳态[159],还可以解析内质网中错误折叠蛋白的积累以及细胞在生理和病理状态下的应激反应,调节内质网中的氧化还原状态调节二硫键的形成,可以优化UPR以提高外源蛋白的表达效率[160],促进蛋白质的正确折叠[161],便于后续纯化.但这些手段在实际应用中可能存在调控不够精确、效果不显著等问题.例如,通过过表达某些分子伴侣来促进蛋白折叠,但过量表达可能会对细胞生理产生负面影响,甚至干扰其他正常蛋白的折叠过程.不同外源蛋白在植物细胞内的折叠和积累情况差异较大,对于一些结构复杂或难以正确折叠的蛋白,优化内质网环境可能无法取得理想的表达效率和质量提升效果.此外,植物细胞的生理状态、环境因素等也会对外源蛋白的表达和折叠产生影响,导致优化效果不稳定. ...
Advances on the Expressions of Foreign Proteins in Plants
1
2023
... 外源蛋白在植物细胞内还可能引起内质网胁迫,影响蛋白的正确折叠和积累,该情况可以通过优化内质网蛋白折叠与质量监控系统和增强内质网相关降解ERAD 途径来处理和清除内质网中错误折叠的有缺陷的蛋白质,可以提高外源蛋白的表达效率[158],维持内质网稳态[159],还可以解析内质网中错误折叠蛋白的积累以及细胞在生理和病理状态下的应激反应,调节内质网中的氧化还原状态调节二硫键的形成,可以优化UPR以提高外源蛋白的表达效率[160],促进蛋白质的正确折叠[161],便于后续纯化.但这些手段在实际应用中可能存在调控不够精确、效果不显著等问题.例如,通过过表达某些分子伴侣来促进蛋白折叠,但过量表达可能会对细胞生理产生负面影响,甚至干扰其他正常蛋白的折叠过程.不同外源蛋白在植物细胞内的折叠和积累情况差异较大,对于一些结构复杂或难以正确折叠的蛋白,优化内质网环境可能无法取得理想的表达效率和质量提升效果.此外,植物细胞的生理状态、环境因素等也会对外源蛋白的表达和折叠产生影响,导致优化效果不稳定. ...
Protein quality control and aggregation in the endoplasmic reticulum: From basic to bedside
1
2023
... 外源蛋白在植物细胞内还可能引起内质网胁迫,影响蛋白的正确折叠和积累,该情况可以通过优化内质网蛋白折叠与质量监控系统和增强内质网相关降解ERAD 途径来处理和清除内质网中错误折叠的有缺陷的蛋白质,可以提高外源蛋白的表达效率[158],维持内质网稳态[159],还可以解析内质网中错误折叠蛋白的积累以及细胞在生理和病理状态下的应激反应,调节内质网中的氧化还原状态调节二硫键的形成,可以优化UPR以提高外源蛋白的表达效率[160],促进蛋白质的正确折叠[161],便于后续纯化.但这些手段在实际应用中可能存在调控不够精确、效果不显著等问题.例如,通过过表达某些分子伴侣来促进蛋白折叠,但过量表达可能会对细胞生理产生负面影响,甚至干扰其他正常蛋白的折叠过程.不同外源蛋白在植物细胞内的折叠和积累情况差异较大,对于一些结构复杂或难以正确折叠的蛋白,优化内质网环境可能无法取得理想的表达效率和质量提升效果.此外,植物细胞的生理状态、环境因素等也会对外源蛋白的表达和折叠产生影响,导致优化效果不稳定. ...
Deciphering ER stress-unfolded protein response relationship by visualizing unfolded proteins in the ER
1
2024
... 外源蛋白在植物细胞内还可能引起内质网胁迫,影响蛋白的正确折叠和积累,该情况可以通过优化内质网蛋白折叠与质量监控系统和增强内质网相关降解ERAD 途径来处理和清除内质网中错误折叠的有缺陷的蛋白质,可以提高外源蛋白的表达效率[158],维持内质网稳态[159],还可以解析内质网中错误折叠蛋白的积累以及细胞在生理和病理状态下的应激反应,调节内质网中的氧化还原状态调节二硫键的形成,可以优化UPR以提高外源蛋白的表达效率[160],促进蛋白质的正确折叠[161],便于后续纯化.但这些手段在实际应用中可能存在调控不够精确、效果不显著等问题.例如,通过过表达某些分子伴侣来促进蛋白折叠,但过量表达可能会对细胞生理产生负面影响,甚至干扰其他正常蛋白的折叠过程.不同外源蛋白在植物细胞内的折叠和积累情况差异较大,对于一些结构复杂或难以正确折叠的蛋白,优化内质网环境可能无法取得理想的表达效率和质量提升效果.此外,植物细胞的生理状态、环境因素等也会对外源蛋白的表达和折叠产生影响,导致优化效果不稳定. ...
Oxidative protein folding fidelity and redoxtasis in the endoplasmic reticulum
1
2022
... 外源蛋白在植物细胞内还可能引起内质网胁迫,影响蛋白的正确折叠和积累,该情况可以通过优化内质网蛋白折叠与质量监控系统和增强内质网相关降解ERAD 途径来处理和清除内质网中错误折叠的有缺陷的蛋白质,可以提高外源蛋白的表达效率[158],维持内质网稳态[159],还可以解析内质网中错误折叠蛋白的积累以及细胞在生理和病理状态下的应激反应,调节内质网中的氧化还原状态调节二硫键的形成,可以优化UPR以提高外源蛋白的表达效率[160],促进蛋白质的正确折叠[161],便于后续纯化.但这些手段在实际应用中可能存在调控不够精确、效果不显著等问题.例如,通过过表达某些分子伴侣来促进蛋白折叠,但过量表达可能会对细胞生理产生负面影响,甚至干扰其他正常蛋白的折叠过程.不同外源蛋白在植物细胞内的折叠和积累情况差异较大,对于一些结构复杂或难以正确折叠的蛋白,优化内质网环境可能无法取得理想的表达效率和质量提升效果.此外,植物细胞的生理状态、环境因素等也会对外源蛋白的表达和折叠产生影响,导致优化效果不稳定. ...
一种植物蛋白纯化用研磨装置
1
2022
... 在物理方面,康跻耀等发明了一种植物蛋白纯化用研磨装置,通过第一研磨槽和第二研磨槽可以对植物蛋白原料进行双层研磨,可以保证研磨质量,从而保证纯化质量,同时该装置为一体式设计,使用方便,效率高[162],唐静秋等公开了一种植物蛋白纯化用固液分离装置,通过液压缸带动压板对植物原料进行挤压,使得水分从浸提箱上的网孔排出滴落在浸提腔中,从而实现固液分离[163].以上方法给植物外源物质的分离与纯化提供了一定的新思路. ...
A Grinding Device for the Purification of Plant Proteins
1
2022
... 在物理方面,康跻耀等发明了一种植物蛋白纯化用研磨装置,通过第一研磨槽和第二研磨槽可以对植物蛋白原料进行双层研磨,可以保证研磨质量,从而保证纯化质量,同时该装置为一体式设计,使用方便,效率高[162],唐静秋等公开了一种植物蛋白纯化用固液分离装置,通过液压缸带动压板对植物原料进行挤压,使得水分从浸提箱上的网孔排出滴落在浸提腔中,从而实现固液分离[163].以上方法给植物外源物质的分离与纯化提供了一定的新思路. ...
一种植物蛋白纯化用固液分离装置
1
2022
... 在物理方面,康跻耀等发明了一种植物蛋白纯化用研磨装置,通过第一研磨槽和第二研磨槽可以对植物蛋白原料进行双层研磨,可以保证研磨质量,从而保证纯化质量,同时该装置为一体式设计,使用方便,效率高[162],唐静秋等公开了一种植物蛋白纯化用固液分离装置,通过液压缸带动压板对植物原料进行挤压,使得水分从浸提箱上的网孔排出滴落在浸提腔中,从而实现固液分离[163].以上方法给植物外源物质的分离与纯化提供了一定的新思路. ...
A Solid-Liquid Separation Device for Plant Protein Purification
1
2022
... 在物理方面,康跻耀等发明了一种植物蛋白纯化用研磨装置,通过第一研磨槽和第二研磨槽可以对植物蛋白原料进行双层研磨,可以保证研磨质量,从而保证纯化质量,同时该装置为一体式设计,使用方便,效率高[162],唐静秋等公开了一种植物蛋白纯化用固液分离装置,通过液压缸带动压板对植物原料进行挤压,使得水分从浸提箱上的网孔排出滴落在浸提腔中,从而实现固液分离[163].以上方法给植物外源物质的分离与纯化提供了一定的新思路. ...
High Level Production of Monoclonal Antibodies Using an Optimized Plant Expression System
1
... 植物相比传统的细胞表达系统有几个潜在的好处.它们的栽培很简单,因为不需要无菌环境,且相对便宜,只需使用常规肥料即可快速生长,而动物细胞培养系统则需要昂贵的培养基,通常还需要高等级的生物安全设施.植物的生产平台具有无限的可扩展性,扩展所需的成本远低于更传统的大肠杆菌与酵母菌发酵表达平台的扩展成本,植物表达系统的生产成本可以低至每千克10-20美元,而酵母表达系统每千克疫苗的生产成本约为200-300美元[164],细菌表达系统的疫苗生产成本约为每千克100-200美元,哺乳动物细胞表达系统每千克疫苗的生产成本约为260美元[165].植物可以在温室或垂直农场中轻松扩大生产,这样可以实现多吨级的产量.快速和准确地生产疫苗和治疗性抗体在应对即将成为全球大流行病之前的威胁至关重要.瞬时表达允许快速生成产品,获得基因序列后大约8周即可完成,而且由于所用植物并未经过基因改造,因此商业生产的监管要求可能较少.这样的快速生产方式操作简便,能够快速应对大型流行病.此外,植物可以进行翻译后修饰(PTM),产生类似于哺乳动物相应物的糖基化产品.通过瞬时共同表达伴侣蛋白或其他酶,可以实现蛋白质的正确折叠和处理[166].由于表达载体开发的现代化,特别是能够瞬时表达的载体,新的植物表达系统还提供了源自哺乳动物细胞培养物的表达技术无法匹配的速度和易处理性[3].“解构的”病毒载体系统如pEAQ、magnICON和双生病毒表达平台的进展已经成功地解决了植物中生物制药生产的蛋白质表达水平、速度和一致性不足等问题[167-169]. ...
Plant Expression Platform for the Production of Recombinant Pharmaceutical Proteins
1
2014
... 植物相比传统的细胞表达系统有几个潜在的好处.它们的栽培很简单,因为不需要无菌环境,且相对便宜,只需使用常规肥料即可快速生长,而动物细胞培养系统则需要昂贵的培养基,通常还需要高等级的生物安全设施.植物的生产平台具有无限的可扩展性,扩展所需的成本远低于更传统的大肠杆菌与酵母菌发酵表达平台的扩展成本,植物表达系统的生产成本可以低至每千克10-20美元,而酵母表达系统每千克疫苗的生产成本约为200-300美元[164],细菌表达系统的疫苗生产成本约为每千克100-200美元,哺乳动物细胞表达系统每千克疫苗的生产成本约为260美元[165].植物可以在温室或垂直农场中轻松扩大生产,这样可以实现多吨级的产量.快速和准确地生产疫苗和治疗性抗体在应对即将成为全球大流行病之前的威胁至关重要.瞬时表达允许快速生成产品,获得基因序列后大约8周即可完成,而且由于所用植物并未经过基因改造,因此商业生产的监管要求可能较少.这样的快速生产方式操作简便,能够快速应对大型流行病.此外,植物可以进行翻译后修饰(PTM),产生类似于哺乳动物相应物的糖基化产品.通过瞬时共同表达伴侣蛋白或其他酶,可以实现蛋白质的正确折叠和处理[166].由于表达载体开发的现代化,特别是能够瞬时表达的载体,新的植物表达系统还提供了源自哺乳动物细胞培养物的表达技术无法匹配的速度和易处理性[3].“解构的”病毒载体系统如pEAQ、magnICON和双生病毒表达平台的进展已经成功地解决了植物中生物制药生产的蛋白质表达水平、速度和一致性不足等问题[167-169]. ...
2
... 植物相比传统的细胞表达系统有几个潜在的好处.它们的栽培很简单,因为不需要无菌环境,且相对便宜,只需使用常规肥料即可快速生长,而动物细胞培养系统则需要昂贵的培养基,通常还需要高等级的生物安全设施.植物的生产平台具有无限的可扩展性,扩展所需的成本远低于更传统的大肠杆菌与酵母菌发酵表达平台的扩展成本,植物表达系统的生产成本可以低至每千克10-20美元,而酵母表达系统每千克疫苗的生产成本约为200-300美元[164],细菌表达系统的疫苗生产成本约为每千克100-200美元,哺乳动物细胞表达系统每千克疫苗的生产成本约为260美元[165].植物可以在温室或垂直农场中轻松扩大生产,这样可以实现多吨级的产量.快速和准确地生产疫苗和治疗性抗体在应对即将成为全球大流行病之前的威胁至关重要.瞬时表达允许快速生成产品,获得基因序列后大约8周即可完成,而且由于所用植物并未经过基因改造,因此商业生产的监管要求可能较少.这样的快速生产方式操作简便,能够快速应对大型流行病.此外,植物可以进行翻译后修饰(PTM),产生类似于哺乳动物相应物的糖基化产品.通过瞬时共同表达伴侣蛋白或其他酶,可以实现蛋白质的正确折叠和处理[166].由于表达载体开发的现代化,特别是能够瞬时表达的载体,新的植物表达系统还提供了源自哺乳动物细胞培养物的表达技术无法匹配的速度和易处理性[3].“解构的”病毒载体系统如pEAQ、magnICON和双生病毒表达平台的进展已经成功地解决了植物中生物制药生产的蛋白质表达水平、速度和一致性不足等问题[167-169]. ...
Other Images/Table from this Article
{kind=link}