|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
CRISPR/Cas systems and their applications in gene editing with filamentous fungi
Synthetic Biology Journal
2024, 5 (3):
672-693.
DOI: 10.12211/2096-8280.2023-097
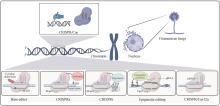
Filamentous fungi, which present distinct morphology and cell structure, play a critical role in human health as well as industrial and agricultural production. However, the unique characteristics of filamentous fungi make them difficult to be manipulated with traditional genetic engineering methods. Thus, the development of an efficient gene editing system is essential for exploring biological resources and understanding metabolic processes in filamentous fungi. The development of the Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR associated protein (CRISPR/Cas) system promotes more efficient and effective gene editing in different species, and brings a revolutionary breakthrough in fungal fundamental research and applications. In this review, we first briefly introduce the history, working mechanism, and classifications of the CRISPR/Cas mediated gene editing system. Next, we comment the functional components of CRISPR/Cas9 such as selective marker, Cas9 and gRNA and the delivery methods of these components in various filamentous fungi. Furthermore, we systematically discuss the applications of CRISPR related technologies, including CRISPR/Cas12, base-editor, CRISPRa, CRISPRi and CRISPR mediated epigenetic regulation, in the genetic engineering of filamentous fungi, particularly in marine-derived filamentous fungi. Finally, we address challenges with relative low gene editing efficiency and off-targets effects in engineering filamentous fungi, and highlight the potential solutions for developing novel CRISPR/Cas-based gene editing systems. This review can provide guidance for developing an efficient gene editing platform in filamentous fungi and pave the way for further exploration of the secondary metabolites and establishment of robust fungal cell factories.
Table 1
Naturally occurring major Cas homologues
Extracts from the Article
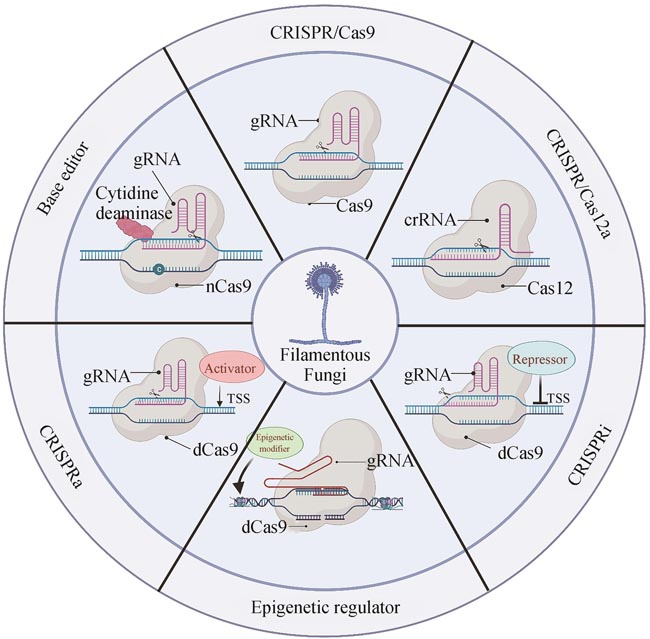
CRISPR/Cas系统主要可分为两大类(ClassⅠ和ClassⅡ),第一大类系统需要多个Cas蛋白形成复合体协同工作,包括TypeⅠ、Ⅲ和Ⅳ型,而第二大类系统仅需一个Cas蛋白就能够切割目标DNA,包括TypeⅡ、Ⅴ和Ⅵ类型[14],这类系统较为简单,研究也更加透彻,是生物技术应用的首选。目前应用比较广泛的CRISPR/Cas,如CRISPR/Cas9、CRISPR/Cas12、CRISPR/Cas13等均为ClassⅡ系统。CRISPR/Cas9因具有简单、快捷等优势,已成为CRISPR/Cas系统中应用最为广泛的技术。来自化脓性链球菌(Streptococcus pyogenes)的Cas9(SpCas9)系统是双RNA引导的——trans-activating crRNA(tracrRNA)靶向DNA,CRISPR associated RNA(crRNA)负责与Cas9形成复合物,并因其识别简单的NGG PAM序列,获得了广泛性应用。此外,研究者们仍一直积极地探索其他具有不同的蛋白分子量、PAM要求和底物偏好的CRISPR系统,仅Cas9的同源蛋白就有70多种已被详细评估过理化活性[15]。在过去的几年里,已经有超过20种不同的Cas同源蛋白被应用于基因编辑,为识别不同PAM序列提供了更多的靶点选择,其中一些典型性代表汇总于表1。
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[13,89]。由于Cas9的持续表达会增加非特异性编辑的风险,同时对一些菌株还可能会产生细胞毒性,而可诱导型启动子能够在基因编辑期间调节CRISPR/Cas9系统的活力,并减少脱靶效应,所以在有特殊需求的情况下,研究者通常会选择可诱导型启动子。常用的可诱导型启动子包括淀粉诱导的启动子amyB、木聚糖诱导的启动子xylA、四环素诱导的启动子tetON等(表3)[73-74,92]。已有研究表明,构建不同丝状真菌的CRISPR/Cas9的基因编辑体系时,要根据菌株特性和实际情况选择最佳Cas9蛋白表达启动子,以保证CRISPR/Cas9系统能够在目标菌株中高效合理地发挥作用。
将催化失活的Cas蛋白与目标DNA的靶向结合形成空间位阻效应可导致基因表达的抑制,若将dCas9与转录阻遏因子融合则能实现高效的转录沉默,这个过程被称为CRISPR干扰(CRISPRi)[131-132]。CRISPRi系统既可结合于目标基因的启动子以阻止RNA聚合酶结合从而抑制转录起始,还可与开放阅读框靶向结合,抑制转录延伸[131,133]。目前,CRISPRi系统已应用于多种模式生物,然而在真菌微生物中,该基因干扰工具主要应用于酵母菌,在丝状真菌中的应用研究极少。2023年,华中农业大学谢卡斌团队[134]分别将dCas9与两个不同的转录阻遏结构域SRDX(SUPERMAN Repression domain)和Mxi1(repressor domain)融合,并测试了这两个体系在植物病原菌中的转录抑制效果。研究发现,在稻瘟病菌中CRISPR/dCas9-MxI1的抑制效果要高于CRISPR/dCas9-SRDX,且由MoRP27启动子驱动的dCas9-SRDX表达体系与MoTEF1启动子相比,可将抑制效率提高上百倍。此外,该研究还揭示gRNA结合于转录起始位点-100 bp到-200 bp之间可起到最佳的转录抑制效果,并利用tRNA-gRNA组装成多gRNA表达体系,在两株不同的植物病原菌中实现多基因高效抑制。相较于shRNA干扰和CRISPR介导的敲除,CRISPRi具有不损伤基因序列、特异性高、抑制效果强和实用性广等优势,在真菌基因功能研究尤其是必需关键基因的功能研究方面,具有巨大应用前景。
表观遗传修饰是指染色体DNA和组蛋白上的化学修饰,包括DNA甲基化、组蛋白甲基化、组蛋白乙酰化、组蛋白磷酸化等,在不改变DNA序列的情况下,这些修饰可以通过改变染色质状态来控制基因的表达,并具有可遗传性。CRISPR表观遗传编辑是指将失去切割活性的Cas蛋白与表观遗传修饰因子融合来靶向改变目标区域的表观遗传修饰标记,从而影响基因的转录[135]。与传统的作用于全基因组的表观遗传调控手段相比,CRISPR表观遗传编辑因其可实现高效的靶向特异性表观遗传修饰,在生命科学研究中具有极大的应用前景。CRISPR表观遗传编辑系统的改造对象主要包括DNA的甲基化修饰和染色质上组蛋白的表观修饰。基于人源E1A相关蛋白p300的组蛋白乙酰转移酶(HAT)核心结构域与dCas9融合的基因激活系统,在人类细胞和多个物种表现出精准且高效的基因诱导活性[136-137]。2021年,研究人员首次将dCas9-p300表观编辑系统应用于丝状真菌中[138]。在黑曲霉中,dCas9-p300可诱导多个次级代谢产物合成基因的表达,并提高了伏马毒素B2的产量。此外,研究人员还将dCas9分别与黑曲霉内源性组蛋白乙酰化酶GcnE、组蛋白去乙酰化酶HosA和RpdA融合,构建了多个CRISPR表观遗传编辑系统。测试结果发现dCas9-GcnE/HosA/RpdA编辑体系均可抑制次级代谢产物合成基breF的表达,dCas9-HosA还可上调色素合成基因fwnA的表达[138]。这些研究结果表明丝状真菌中还存在其他因素可影响内源性表观遗传调控因子的调控效果,在推广应用丝状真菌来源的各种表观调控因子之前,还需更深入地研究这些表观调控因子的作用机制。
CRISPRoff/on是另外一个划时代的表观遗传调控系统,是由美国加州大学旧金山分校和麻省理工学院的研究团队联合开发[139]。CRISPRoff系统将转录抑制因子KRAB和DNA甲基转移酶D3A-D3L分别融合到dCas9的N末端/C末端,能够高特异性地持续沉默基因表达。为了逆转CRISPRoff系统介导的基因沉默,该团队还利用DNA去甲基化酶TET1与dCas9融合,并结合MS2-MCP转录激活系统,开发出一种可介导靶位点DNA去甲基化和基因转录激活的新工具CRISPRon系统,该系统可有效逆转人源细胞中由CRISPRoff介导的DNA甲基化修饰和转录抑制。近期,荷兰乌得勒支大学的研究人员首次将CRISPRoff/on应用于黑曲霉中[140],CRISPRoff系统对黑曲霉中的flbA基因的抑制效率高达100%,即便在去除CRISPRoff编辑质粒之后,仍表现出可持续和可传代的抑制效果,且这些抑制效果可被导入的CRISPRon编辑质粒高效逆转。
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
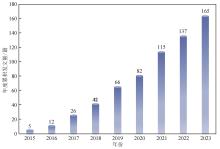
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Application of CRISPR in filamentous fungi and macrofungi: from component function to development potentiality 5 2023 ... CRISPR/Cas9系统介导的丝状真菌基因编辑技术始于2015年[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Development of the CRISPR/Cas9 system for targeted gene disruption in Aspergillus fumigatus 4 2015 ... A. niger[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Development of a genome-editing CRISPR/Cas9 system in thermophilic fungal Myceliophthora species and its application to hyper-cellulase production strain engineering 3 2017 ... M. thermophila[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Forced recycling of an AMA1-based genome-editing plasmid allows for efficient multiple gene deletion/integration in the industrial filamentous fungus Aspergillus oryzae 6 2019 ... M. thermophila[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ 5S rRNA promoter for guide RNA expression enabled highly efficient CRISPR/Cas9 genome editing in Aspergillus niger 5 2019 ... A. niger[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Development of versatile and efficient genetic tools for the marine-derived fungus Aspergillus terreus RA2905 3 2022 ... M. thermophila[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Highly efficient single base editing in Aspergillus niger with CRISPR/Cas9 cytidine deaminase fusion 2 2019 ... M. thermophila[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Development of a novel strategy for fungal transformation based on a mutant locus conferring carboxin-resistance in Magnaporthe oryzae 2 2016 ... 抗性基因是发展最早、应用最广的一类筛选标记,具有应用方便、选择效率高、功能稳定等特点.抗性基因编码可使抗生素、除草剂等药物失活的酶(如潮霉素磷酸转移酶),从而使导入抗性基因的重组细胞可以在含有相应药物的培养基中生长.目前丝状真菌可使用的抗生素主要有:潮霉素(hygromycin)、遗传霉素(geneticin)、博来霉素(bleomycin)、吡啶硫胺素(pyrithiamine)、草铵膦(glufosinate)和萎锈灵(carboxin)等[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Applications of CRISPR/Cas9 in the synthesis of secondary metabolites in filamentous fungi 1 2021 ... 体内表达体系是将密码子优化过的Cas9蛋白表达序列置于真菌适用的启动子后面,并将组装好的DNA序列导入真菌细胞中在体内进行表达.启动子驱动表达能力的强弱是影响外源性Cas9蛋白表达效率的重要因素,因此选择合适的启动子对CRISPR/Cas9系统的功能发挥具有重要意义.用于Cas9表达的启动子为RNA聚合酶Ⅱ启动子,其主要分为组成型启动子和诱导型启动子.由于组成型启动子驱动基因表达不受空间和时间因素的影响,可在真菌细胞稳定持续表达,大多数研究中会优先选用其来驱动Cas9蛋白在丝状真菌中的表达.丝状真菌CRISPR/Cas9系统常用的组成型启动子主要是真菌来源的启动子序列,如tef1启动子、gpdA启动子和trpC启动子[ High-level expression of highly active and thermostable trehalase from Myceliophthora thermophila in Aspergillus niger by using the CRISPR/Cas9 tool and its application in ethanol fermentation 1 2020 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Development of a versatile and conventional technique for gene disruption in filamentous fungi based on CRISPR-Cas9 technology 2 2017 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Deletion of a rare fungal PKS CgPKS11 promotes chaetoglobosin A biosynthesis, yet defers the growth and development of Chaetomium globosum 1 2021 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ The Ustilago hordei-barley interaction is a versatile system for characterization of fungal effectors 1 2021 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Efficient genome editing using tRNA promoter-driven CRISPR/Cas9 gRNA in Aspergillus niger 1 2018 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Genome editing in Shiraia bambusicola using CRISPR-Cas9 system 2 2017 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Functional reconstitution of a fungal natural product gene cluster by advanced genome editing 2 2017 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ CRISPR/Cas9-mediated genome editing in Penicillium oxalicum and Trichoderma reesei using 5S rRNA promoter-driven guide RNAs 2 2021 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Identification and characterization of the determinants of copper resistance in the acidophilic fungus Acidomyces richmondensis MEY-1 using the CRISPR/Cas9 system 1 2023 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ A versatile in vivo DNA assembly toolbox for fungal strain engineering 1 2022 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Genome editing in Penicillium chrysogenum using Cas9 ribonucleoprotein particles 1 2018 ... Promoters for expressing Cas9/gRNA in filamentous fungi
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Production of L-malic acid by metabolically engineered Aspergillus nidulans based on efficient CRISPR-Cas9 and cre-loxP systems 1 2023 ... gRNA的高效表达也是CRISPR/Cas9系统发挥作用的关键因素.与Cas9的表达一致,gRNA的表达也分为体内和体外表达.gRNA的体外表达通常利用体外转录试剂盒由T7 RNA聚合酶转录而成[ CRISPR/Cas9基因编辑技术在丝状真菌中的应用 1 2022 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Research progress on application of CRISPR/Cas9 gene editing technique in filamentous fungi 1 2022 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Development of an efficient vector system for gene knock-out and near in-cis gene complementation in the sugarcane smut fungus 1 2017 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Generation of Trichoderma harzianum with pyr4 auxotrophic marker by using the CRISPR/Cas9 system 1 2021 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ CRISPR-Cas9 induces point mutation in the mucormycosis fungus Rhizopus delemar 1 2019 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Methods for genetic transformation of filamentous fungi 1 2017 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Kinetic basis for DNA target specificity of CRISPR-Cas12a 1 2018 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[ Multiplexed genome engineering by Cas12a and CRISPR arrays encoded on single transcripts 1 2019 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[ Multiplex genome editing in Ashbya gossypii using CRISPR-Cpf1 2 2020 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[
Other Images/Table from this Article
|