|
||||||||||||||||||||||||||||||||||||||||||||||||
|
CRISPR/Cas systems and their applications in gene editing with filamentous fungi
Synthetic Biology Journal
2024, 5 (3):
672-693.
DOI: 10.12211/2096-8280.2023-097
Filamentous fungi, which present distinct morphology and cell structure, play a critical role in human health as well as industrial and agricultural production. However, the unique characteristics of filamentous fungi make them difficult to be manipulated with traditional genetic engineering methods. Thus, the development of an efficient gene editing system is essential for exploring biological resources and understanding metabolic processes in filamentous fungi. The development of the Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR associated protein (CRISPR/Cas) system promotes more efficient and effective gene editing in different species, and brings a revolutionary breakthrough in fungal fundamental research and applications. In this review, we first briefly introduce the history, working mechanism, and classifications of the CRISPR/Cas mediated gene editing system. Next, we comment the functional components of CRISPR/Cas9 such as selective marker, Cas9 and gRNA and the delivery methods of these components in various filamentous fungi. Furthermore, we systematically discuss the applications of CRISPR related technologies, including CRISPR/Cas12, base-editor, CRISPRa, CRISPRi and CRISPR mediated epigenetic regulation, in the genetic engineering of filamentous fungi, particularly in marine-derived filamentous fungi. Finally, we address challenges with relative low gene editing efficiency and off-targets effects in engineering filamentous fungi, and highlight the potential solutions for developing novel CRISPR/Cas-based gene editing systems. This review can provide guidance for developing an efficient gene editing platform in filamentous fungi and pave the way for further exploration of the secondary metabolites and establishment of robust fungal cell factories.
Table 4
Examples of the CRISPR/Cas12a system-assisted gene editing in filamentous fungi
Extracts from the Article
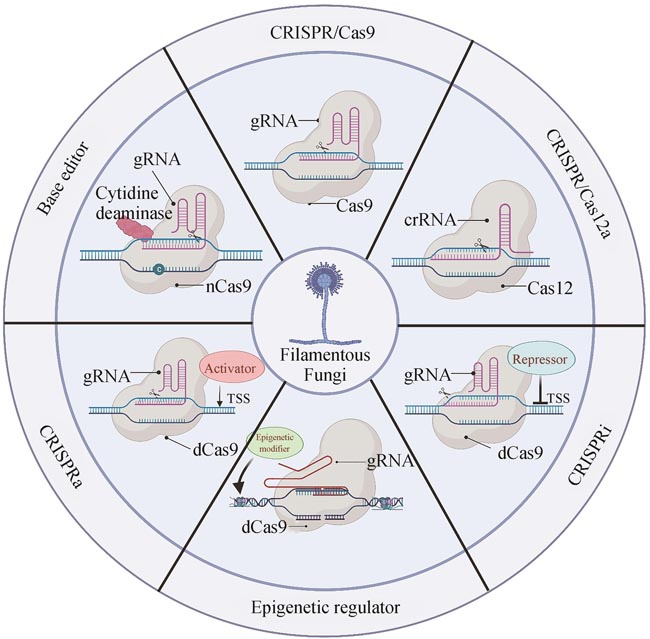
Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[25],Cas12a的识别序列(5′-TTTV-3′)和切割方式(切割DNA双链产生黏性末端)与Cas9有较大的差异,因此,CRISPR/Cas12a作为进阶版基因编辑工具可扩充CRISPR/Cas系统及其衍生技术体系的基因编辑范围[38]。在编辑效率方面,Cas12a的编辑效率要低于Cas9,但Cas12a具有更低的容错率和更高的特异性[104]。Cas12a比较重要的一个特点是其只需单一RNA引导,且Cas12a自带的RNA酶结构域可处理preCrRNAs产生成熟CrRNAs,利用该特性,Cas12a系统则可以在一个质粒上仅用一个启动子串联多个CrRNA,由Cas12a剪切合成多个单独的成熟CrRNA,为多基因编辑提供便利[105]。基于CRISPR/Cas12a的多基因编辑系统,中国科学院天津工业生物技术研究所田朝光团队[44]在嗜热毁丝菌(M. thermophila)成功实现高效的多基因编辑,并进一步开发了CRISPR/Cas12辅助的标记回收技术。类似地,Jimenez等[106]利用CRISPR/Cas12a多基因编辑系统在阿舒氏囊霉(A. gossypii)中敲除了5个营养缺陷标记基因。Cas12a还具有PAM特异性识别和对单链DNA的非特异性切割的独特属性,在DNA诊断方面拥有巨大潜力[107]。基于此,研究人员将PCR扩增技术与CRISPR/Cas12a相结合开发了一种检测系统,可用于监测小麦中禾谷镰刀菌(F. graminearum)侵染情况[108]。此外,研究者们还在构巢曲霉(A. nidulans)、棘孢曲霉(A. aculeatus)等丝状真菌中建立了基于CRISPR/Cas12a的基因编辑工具,具体信息详见表4。CRISPR/Cas12a和CRISPR/Cas9基因编辑技术相互补充,研究者可根据实际需求选择最适合的编辑工具,保障丝状真菌的基因编辑工作高效进行。
胞嘧啶碱基编辑器是将nCas9(nickase Cas9)与胞嘧啶脱氨酶和尿嘧啶糖基化酶抑制子融合,gRNA引导融合蛋白到靶位点时,暴露出来的非靶向DNA单链上的胞嘧啶C可被脱氨酶转换为尿嘧啶U,而U与胸腺嘧啶T的具有相同的碱基配对规则,在DNA修复时,尿嘧啶糖基化酶抑制子会抑制U→C的修复,提高U转换为T的修复效率,从而完成C→T的碱基转换[60-63]。2019年,华南理工大学潘力团队[58]首次将胞嘧啶碱基编辑器应用于黑曲霉中,其编辑效率可达47.36%~100%,且胞嘧啶碱基编辑器在黑曲霉中的靶向编辑窗口略大于其他物种。随后,中国科学院天津工业生物技术研究所田朝光团队[123]又进一步在嗜热毁丝霉中构建了三个不同类型的胞嘧啶碱基编辑器(Mtevo-BE4max、MtGAM-BE4max和Mtevo-CDA1)。测试结果显示,由海七鳃鳗胞嘧啶脱氨酶融合而成的胞嘧啶碱基编辑器(Mtevo-CDA1)在嗜热毁丝霉中编辑效率要高于另外两个由大鼠嘧啶脱氨酶融合而成的编辑器(Mtevo-BE4max和MtGAM-BE4max)。研究人员还利用Mtevo-CDA1深入研究了嗜热毁丝霉中转录调控因子Mtclr-2的功能,发现该蛋白的DNA结合结构域对培养基中纤维素的响应起关键作用,而真菌特异性结构域则与菌丝生长存在密切关系。该研究说明碱基编辑器可为研究丝状真菌中蛋白特殊结构域功能提供另一种高效的编辑工具。近期,美国莱斯大学的高雪团队[124]在丝状真菌中开发了基于胞嘧啶单碱基编辑器的多位点基因编辑系统,该体系利用CBE对功能基因中密码子CAG、CAA、CGA以及反义链中的CCA进行C→T的碱基转换,将终止密码子引入基因序列,致使基因失活,并结合tRNA-gRNA阵列表达体系,实现多个基因的同时失活。该研究利用CBE编辑系统对构巢曲霉中的表观遗传因子进行组合灭活,揭示了多种表观遗传调控因子在真菌次级代谢中的协同调控作用,还筛选到4种新型的天然产物。基于CBE的多位点基因编辑系统显著提高了丝状真菌基因组操作的能力,同时也为挖掘新型真菌活性天然产物提供了新的思路和策略。
在部分真菌中也可直接使用哺乳动物细胞中的表达系统,如在竹黄菌(S. bambusicola)中,研究者则直接利用人延长因子1α启动子来驱动Cas9蛋白的表达[ Production of L-malic acid by metabolically engineered Aspergillus nidulans based on efficient CRISPR-Cas9 and cre-loxP systems 1 2023 ... gRNA的高效表达也是CRISPR/Cas9系统发挥作用的关键因素.与Cas9的表达一致,gRNA的表达也分为体内和体外表达.gRNA的体外表达通常利用体外转录试剂盒由T7 RNA聚合酶转录而成[ CRISPR/Cas9基因编辑技术在丝状真菌中的应用 1 2022 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Research progress on application of CRISPR/Cas9 gene editing technique in filamentous fungi 1 2022 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Development of an efficient vector system for gene knock-out and near in-cis gene complementation in the sugarcane smut fungus 1 2017 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Generation of Trichoderma harzianum with pyr4 auxotrophic marker by using the CRISPR/Cas9 system 1 2021 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ CRISPR-Cas9 induces point mutation in the mucormycosis fungus Rhizopus delemar 1 2019 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Methods for genetic transformation of filamentous fungi 1 2017 ... 将外源组装的Cas9蛋白与gRNA表达质粒(DNA)、体外转录的gRNA(RNA)或RNP蛋白导入真菌细胞中是CRISPR/Cas9系统高效编辑的另一关键步骤.CRISPR/Cas9系统转化到真菌细胞中的方法有聚乙二醇介导的转化(PMT)、农杆菌 (Agrobacterium tumefaciens)介导的转化(AMT)、电穿孔转化和基因枪转化法[ Kinetic basis for DNA target specificity of CRISPR-Cas12a 1 2018 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[ Multiplexed genome engineering by Cas12a and CRISPR arrays encoded on single transcripts 1 2019 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[ Multiplex genome editing in Ashbya gossypii using CRISPR-Cpf1 2 2020 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[
构巢曲霉 ... CRISPR-Cas12a-assisted nucleic acid detection 1 2018 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[ CRISPR-Cas12a-based diagnostics of wheat fungal diseases 1 2022 ... Cas12a核酸内切酶功能由张锋实验室在2015年首次鉴定[ CRISPR-Cas12a induced DNA double-strand breaks are repaired by multiple pathways with different mutation profiles in Magnaporthe oryzae 1 2022 ... (M. oryzae )
碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage 1 2016 ... 碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ Base editing: advances and therapeutic opportunities 0 2020 Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C: G-to-T: a base editors with higher efficiency and product purity 1 2017 ... 碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ 碱基编辑技术及其在微生物合成生物学中的应用 1 2023 ... 碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ Base editing technology and its application in microbial synthetic biology 1 2023 ... 碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ CRISPR/Cas9 genome editing to demonstrate the contribution of Cyp51A Gly138Ser to azole resistance in Aspergillus fumigatus 1 2018 ... 碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ Genome-wide target specificities of CRISPR RNA-guided programmable deaminases 1 2017 ... 碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ CRISPR-Cas9 base editors and their current role in human therapeutics 1 2023 ... 碱基编辑器是在CRISPR/Cas系统基础上,将突变型Cas与脱氨酶等其他功能蛋白融合,实现不同碱基的转换,最初是由David Liu团队在2016年 开发[ Development of an efficient C-to-T base-editing system and its application to cellulase transcription factor precise engineering in thermophilic fungus Myceliophthora thermophila 1 2022 ... 胞嘧啶碱基编辑器是将nCas9(nickase Cas9)与胞嘧啶脱氨酶和尿嘧啶糖基化酶抑制子融合,gRNA引导融合蛋白到靶位点时,暴露出来的非靶向DNA单链上的胞嘧啶C可被脱氨酶转换为尿嘧啶U,而U与胸腺嘧啶T的具有相同的碱基配对规则,在DNA修复时,尿嘧啶糖基化酶抑制子会抑制U→C的修复,提高U转换为T的修复效率,从而完成C→T的碱基转换[ Multiplex base-editing enables combinatorial epigenetic regulation for genome mining of fungal natural products 1 2023 ... 胞嘧啶碱基编辑器是将nCas9(nickase Cas9)与胞嘧啶脱氨酶和尿嘧啶糖基化酶抑制子融合,gRNA引导融合蛋白到靶位点时,暴露出来的非靶向DNA单链上的胞嘧啶C可被脱氨酶转换为尿嘧啶U,而U与胸腺嘧啶T的具有相同的碱基配对规则,在DNA修复时,尿嘧啶糖基化酶抑制子会抑制U→C的修复,提高U转换为T的修复效率,从而完成C→T的碱基转换[ CRISPR technology: a decade of genome editing is only the beginning 1 2023 ... CRISPR/Cas除了可用上述的基因编辑功能来灭活各种调控因子或替换启动子进而影响基因的表达,还可将丧失切割活性但仍具有RNA引导下与DNA结合能力的Cas蛋白突变体(dCas9/12)与各种效应因子融合来实现位点特异性基因调控[ Modular synthetic biology toolkit for filamentous fungi 1 2021 ... CRISPR介导的转录激活(CRISPRa)主要是通过将核酸内切酶失活的dCas9/12与转录激活因子融合,并在gRNA指导下结合到目标基因的启动子区域,从而增强特定基因的表达[ Highly efficient Cas9-mediated transcriptional programming 2 2015 ... CRISPR介导的转录激活(CRISPRa)主要是通过将核酸内切酶失活的dCas9/12与转录激活因子融合,并在gRNA指导下结合到目标基因的启动子区域,从而增强特定基因的表达[
Other Images/Table from this Article
|