| 领域 | 具体应用 | 检测靶标 | 传感基因线路元件 | 检测限 | 工作范围 | 参考文献 |
|---|
| 环境 监测 | 重金属离子 | As3+ | 转录因子ArsR、转录信号放大器 | 0.1 ppb | 0.1-5 ppb | [32] | | Cd2+ | 转录因子CadR | 0.39 μg/L | 0-60 μg/L | [10] | | As3+ | II型CRISPR系统 | | 0-32 μM | [17] | | U6+ | 双组分系统UzcRS-UrpRS、AND gate、天然系统活性放大器 | 1 μM | 1-5.2 μM | [33] | | Hg2+ | toehold开关、转录因子MerR | 5 nM | 5-7.5 nM | [34] | | Au3+ | 转录因子HspR、重组酶系统 | 5 μM | 5-100 μM | [35] | | Cu2+ | P CopA 启动子 | | 0-50 mM | [36] | | 爆炸残留物 | 2,4-DNT | yqjF启动子 | 4.8 mg/L | 4.8-25 mg/L | [37] | | TNT | TNT核糖开关,记忆开关 | 25 μM | | [38] | | 农药 | 2-苯基苯酚 | 转录因子HbpR、转录信号放大器 | 1 μM | 1-50 μM | [39] | | 有机污染物 | 单环芳烃 | 双组分系统TodTS | 0.04 mg/L | 0.04-1 mg/L | [40] | | 甲醇 | 双组分系统MxcQZ-OmpR | | 0-0.05% | [41] | | 环境病原 细菌 | AHL | 转录因子QscR | 0.01 μM | 0.01-5 μM | [42] | | 医学 诊疗 | 监测体液 物质 | 孕酮 | 从头设计转录因子DLA | 0.16 μg/L | 0.16- 60 μg/L | [43] | | 人血清中 的锌 | 转录因子ZntR、Zur | | 0-20 μM | [44] | | 大麻素类 化合物 | 基于CB2受体的酵母GPCR | 1 nM | | [45] | | 检测疾病标志物以诊断疾病 | 转化生长 因子-β | 双组分系统Smad | 0.024 ng/mL | 0.024- 6.25 ng/mL | [15] | | 胆盐 | 人工跨膜转录因子CadC-TcpP | 28.3 μM | 28.3- 58.99 μM | [46] | | 血红素 | 转录因子HrtR,拨动开关 | 0.12 ppm | | [47] | | 肠炎标志物NO | 转录因子NorR,重组酶记忆模块 | 30 μM | | [48] | | RNA | CRISPR-Cas13a/C2c2系统 | | | [49] | | DNA | CRISPR-Cas12a系统 | | | [50] | | 传感并治疗疾病 | 血液脂肪酸 | 人工转录因子LSR | 5 μM | 5-100 μM | [51] | | 霍乱弧菌群体感应信号CAI-1 | 人工转录因子HR | 细胞密度108 CFU/ml | | [52] | | 原儿茶酸 | 转录因子 PcaV | | 0-1000 μM | [53] | | 硫代硫酸盐 | 双组分系统 ThsRS | 0.016 mM | 0.016-1 mM | [54] | | 阿司匹林 | 复合转录因子Myr-NPR1/NPR4-VanR-VP16 | | 10-250 μM | [55] | | 硝酸甘油 | 级联生化反应和转录因子CREB | 75 µM | | [56] | | 调节肠道 生态 | 鼠李糖、硫酸软骨素、IPTG | 转录因子RhaR和LacI、双组分系统BT3334- BT0267、 CRISPR记忆模块 | 0.3 mM; 0.01 mM; 6 μM | | [57] | | 生物 制造 | 检测目标产物产量辅助菌株筛选 | L-赖氨酸 | 转录因子LysG | 40 mM | 40-320 mM | [12] | | L-半胱氨酸 | 转录因子CcdR | | 0-50 mM | [58] | | 苹果酸 | 转录因子MalR | 5 g/L | 5-15 g/L | [59] | | 代谢动态 调控 | 半乳糖醛酸 | 转录因子ExuR | | 1-100 mg/L | [60] | | 葡萄糖 | 转录因子Mlc | | | [14] | | L-赖氨酸 | 转录因子LysG | | 0.05-8 mM | [61] | | 长链脂肪酸 | 转录因子FadR、TetR | | | [62] | | 优化发酵 参数 | L-乳酸、D-乳酸 | 转录因子EcLldR、PfPdhR | | 15-200 mM;0-50 mM | [13] | | 辅助酶的定向进化 | 乳果糖 | 转录因子LacI-L5 | 5 μM | 10-500 μM | [63] | | 茶碱 | 茶碱核糖开关 | 10 μM | 10-1000 μM | [64] | | 生物碱 | 转录因子RamR | | 1-100 μM | [65] | | 4'-O-甲基去甲酰胺 | 转录因子RamR | 2.5 μM | 2.5-100 μM | [66] | | 食品 安全 | 检测食物毒性物质 | 腐胺 | 转录因子PuuR | 5.37 mM | | [67] | | 毒素黄素 | 转录因子ToxR | 50 nM | 50-500 nM | [68] | | 组胺 | 转录因子HinK | 0.39 ppm | 0.28-18 ppm | [69] | | 四环素 | 转录因子TetR、聚合酶链回收(PSR)放大线路 | 12 ppb | | [70] | | 食品质量 控制 | 柚皮素 | 柚皮素核糖开关 | | 0-0.6 mM | [71] | | 细胞 调控 | 调控细胞 分裂与运动 | 光照 | 转录因子LexRO、基因逻辑门 | 0.059 mW/cm2 | | [72] |
Table 1
Typical genetic circuit-enabled synthetic biosensors and applications
Extracts from the Article
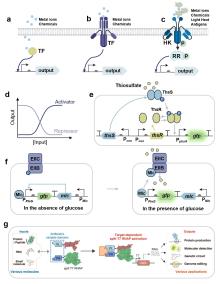
在合成生物学技术体系成熟之前,生物传感器的开发主要依赖于微生物对环境变化应激响应机制的随机发现。这使传感器的开发具有显著的偶然性特征,其技术路径难以实现标准化复现与规模化推广[24]。随着合成基因线路工程化的发展,研究者正逐步建立起标准化、模块化、正交化的生物传感器理性设计策略[7]。该策略的核心在于解析细菌进化形成的天然遗传调控网络(如反馈回路、级联放大、开关切换等动态调控系统)[5,25],从中提取功能明确的基因元件,并通过合成生物学手段将其重构于不同底盘细胞中,形成具有可编程特性的传感系统。从系统架构视角分析,此类传感器可解耦为三个功能单元:传感模块(靶标识别与信号捕获)、处理模块(信号转导与逻辑运算)及输出模块(响应信号生成),三者通过正交化基因线路实现精准耦合[26-30]。当靶标分子触发传感模块时,传感器通过核酸或蛋白质构象变化完成信号转换,然后经处理模块的信息整合,最终激活荧光报告、电化学响应或代谢产物释放等输出,实现对靶标分子的实时动态监测。这种模块化架构赋予传感器高度的设计灵活性,目前已广泛应用于环境监测、生物制造与医学诊断等领域[31],其代表性研究如表1所示。
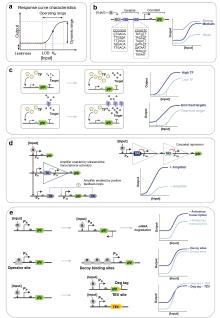
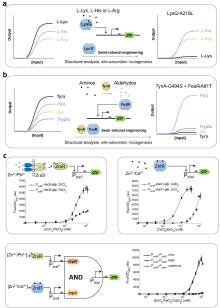
目前已存在众多利用变构转录因子开发的传感器。(1)在生物制造领域,Ding等[14]利用转录抑制因子Mlc和磷酸转移酶组分EIIBC的相互作用构建了一种能监测葡萄糖摄取率的生物传感器(GURBs)。当葡萄糖缺乏时,Mlc结合到P PtsG 启动子上抑制转录;当葡萄糖存在且细胞经EIIBC摄取葡萄糖时,EIIB去磷酸化并招募Mlc,迫使Mlc解除对下游报告基因表达的抑制,从而监测葡萄糖摄取率(图1f)。他们继续利用GURBs在细胞工厂动态调控中央代谢途径的代谢流,成功实现了副产物的减少和目标产物的产量提高。(2)在医学诊疗领域,抑制型转录因子PcaV在结合原儿茶酸后将发生构象变化解除对下游基因表达的抑制,Jiang等[53]基于这一原理在益生菌E. coli Nissle 1917(EcN)中构建了原儿茶酸传感系统,可用于活体疗法或药物递送。(3)在农业科学领域,Haskett等[83]通过整合根瘤菌信号分子scyllo-inosamine合成途径与工程菌感知系统,构建了植物-微生物互作固氮调控网络,感知到信号分子后,工程菌中的转录因子MocR将驱动下游固氮酶相关基因的表达。
尽管已存在上述在胞内作用的变构转录因子机制,跨膜信号转导系统的开发为胞外环境感知提供了新范式[86]。以ToxR家族为代表的单组分系统通过膜整合结构域直接感应外界信号并利用胞内DNA结合域调控基因表达(图1b)[87-88],其中大肠杆菌跨膜转录因子CadABC模块的分子机制已经得到广泛研究[89]:CadC通过周质结构域感知环境pH变化,激活CadA/CadB基因表达[90-91]。Chang等[46]基于模块化设计理念构建了“EMeRALD”合成受体平台,通过将不同感应模块(如胆盐结合域TcpP)与CadC的DNA结合域融合,提出了一种通过更换目的感应模块来实现对不同的目标物进行检测的方法,并实现了咖啡因等分子的特异性检测[92];Shen等[93]对CadC及其结合位点CadO进行定向进化,有效提高镉传感器的性能,建立了一个简便快速的镉离子检测智能手机平台。
总的来说,基于转录因子的传感器具有极其丰富的传感元件结构多样性和广泛的潜在可检测分子库,尽管已经得到了广泛的研究,但其作为生物传感器的潜力仍未得到充分挖掘。目前,其设计与性能调控策略也主要依赖于反复的迭代优化,需建立具备普适性的基因元件调控体系,加强设计与优化的可预测性。此外,机器学习与传感器输出的深度耦合将成为未来的重要发展方向:例如Li等[94]构建了包含4种传感器的全细胞阵列,结合线性回归、反向传播神经网络及最小二乘支持向量机算法,建立了小麦霉菌污染判别模型;Saltepe等[35]通过深度神经网络解析金离子传感器的荧光强度-浓度非线性关系,在短时间内能够实现靶标浓度的精确定量。这些案例展示了人工智能在提升传感器性能与数据分析效率中的关键作用,提供了智能化的信号解读工具。值得注意的是,Komatsu等[95]将人为拆分的T7 RNA聚合酶分别与抗体的重链、轻链可变结构域(VH和VL)融合,构建了一种靶标依赖性RNA聚合酶(TdRNAP),当融合的VH和VL结构域与特定分子靶标相互作用时,拆分的T7 RNA聚合酶将被组装成完整的活化RNA聚合酶,从而驱动T7启动子下游目标基因的表达产生各种传感输出(图1g)。这一通过融合转录调控蛋白与抗体结构域以实现基因线路传感输出的创新方法给转录因子传感器的设计带来了启发,未来可尝试将拆分的转录因子与抗体或其他特异性识别分子融合构建新型人工转录因子。
合成生物传感器的本底泄漏抑制是提升检测敏感性的关键环节,当前研究通过分子互作机制与基因线路拓扑重构等路径实现泄漏的精准调控。在分子调控层面,融合降解标签[44]并诱导表达特异性蛋白酶可靶向降低本底蛋白表达[32],而反向启动子介导的反义RNA转录能通过阻碍核糖体结合或触发RNA降解通路实现翻译级泄漏抑制[184](图8e)。Ho等[185]将分裂内含肽与拆分的目标蛋白分别偶联,并用两个独立启动子控制,使得单一启动子泄漏不足以产生活性蛋白,这种基于AND逻辑的策略成功降低了目标蛋白泄漏表达,在传感器泄漏优化方面有着应用潜力。本文1.4部分提及的断裂内含子反式剪接(SENTR)技术[122]同样有望通过类似策略降低传感器泄漏。在线路设计层面,Wan等[186]通过引入多拷贝的转录因子结合位点作为诱饵(DNA海绵[187])竞争性捕获游离转录因子,使大肠杆菌报告系统泄漏表达降低20倍以上,动态范围提高70倍,同时降低宿主细胞毒性;结合位点空间排布的优化(如调整其与转录起始位点间距[180,188]或在下游增设额外结合位点[189])也可从转录层面有效降低泄漏表达(图8e)。此外,通过正交调控模块的协同级联调控可实现泄漏的精准控制,如Greco等[190]开发的多级控制器(MLC)在转录因子与翻译调节因子同时激活时才能启动报告基因表达,使泄漏水平较传统系统明显降低,动态范围提高1000倍以上。
Other Images/Table from this Article
|