|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Research progress and development trends in the biosynthesis of neutral core human milk oligosaccharides
Synthetic Biology Journal
2025, 6 (5):
1126-1144.
DOI: 10.12211/2096-8280.2025-083
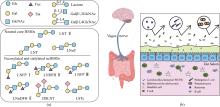
Human milk oligosaccharides (HMOs) are essentially functional and nutritional components found in human milk. They can be primarily classified into fucosylated, neutral core, and sialylated HMOs. Lacto-N-triose Ⅱ (LNT Ⅱ), lacto-N-neotetraose (LNnT), and lacto-N-tetraose (LNT) are common neutral core human milk oligosaccharides (ncHMOs), which can be extended to form longer-chain HMOs and play important roles in intestinal health. In recent years, the biosynthesis of ncHMOs has developed rapidly, and industrial-scale production is from theoretical possibility to practical reality. The synthesis approaches for ncHMOs include chemical synthesis, enzymatic synthesis, and microbial cell synthesis. As the rapid development in biotechnology, enzymatic and microbial cell synthesis have emerged as prominent methods in ncHMOs biosynthesis. Enzymatic synthesis is highly efficient, regioselective, and stereoselective. Currently, glycosyltransferases and glycoside hydrolases represent the two major types of enzymes used for biosynthesizing ncHMOs. Glycosidase-based enzymatic synthesis has demonstrated high conversion rates for LNT Ⅱ and LNnT production. However, the enzymatic synthesis of LNT is less efficient and requires further improvement. Notably, the production of LNnT and LNT typically relies on LNT Ⅱ as a key precursor, requiring a multi-step synthetic strategy. Microbial cell synthesis employs metabolic engineering to construct continuously synthetic pathways in microbial cells such as Escherichia coli and Bacillus subtilis. Knocking out genes in competitive pathway, optimizing genes expression, regenerating cofactors have significantly enhanced the yields of ncHMOs. The biosynthesis of ncHMOs faces several critical challenges, including the low activity and poor substrate specificity of key glycosyltransferases, such as β-1,3-N-acetylglucosaminyltransferase and β-1,3-galactosyltransferase. Additionally, the transporters of LNT Ⅱ and LNnT are not clear in microbial cell. Furthermore, the yields of LNT Ⅱ should be substantially improved for industrial-scale production. Thus, it is important to overcome the interconnected limitations in enzyme engineering (particularly glycosyltransferase specificity and activity), microbial cell modification (focusing on metabolic compatibility and pathway design), and bioprocess optimization (through rational pathway redesign) via an integrated synthetic biology and fermentation engineering approach in the future. These strategies are essential for achieving efficient, cost-effective biosynthesis of ncHMO at industrial scale.
Table 3
Summary of LNnT synthesis by β-1,4-galactosyltransferases and β-galactosidases
Extracts from the Article
酶法合成LNnT主要利用β-1,4-半乳糖基转移酶(β-1,4-galactosyltransferase,β-1,4-GalT,EC 2.4.1.22)和β-半乳糖苷酶(β-galactosidase,EC 3.2.1.23)。β-1,4-GalT能够催化UDP-半乳糖(UDP-galactose,UDP-Gal)的半乳糖基转移至LNT Ⅱ,形成LNnT。Wakarchuk等[54]首次表征了脑膜炎奈瑟氏菌(Neisseria meningitidis)的Nm-LgtB蛋白功能,证实其能够严格识别β-GlcNAc并作为一种β-1,4-半乳糖转移酶催化LNnT合成。酶法合成过程中,不同β-1,4-GalT催化合成LNnT的转化率不同(表3)。
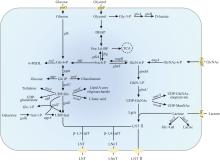
由于大肠杆菌缺乏β-1,3-N-乙酰氨基葡萄糖基转移酶,需将其引入才能利用外源添加的乳糖或N-乙酰氨基葡萄糖和内源合成的UDP-N-乙酰氨基葡萄糖(UDP-GlcNAc)合成LNT Ⅱ(图2)。内源UDP-GlcNAc合成通路包含glmM、glmU以及glmS等关键基因,Zhu等[76]利用不同拷贝数质粒微调UDP-GlcNAc合成通路表达强度,并定点突变了glmS以解除前体物GlcN-6-P的反馈抑制,敲除竞争路径中wecB和nagB后,5 L发酵罐中LNT Ⅱ产量高达46.2 g/L。乳糖作为合成LNT Ⅱ的直接底物,不同大肠杆菌菌株利用其效果不同。与大肠杆菌BL21(DE3)相比,大肠杆菌JM109需过表达lacY以提高乳糖渗透率[77]。此外,还需敲除lacZ防止乳糖降解[77]。Hu等[78]利用几丁质降解产物GlcNAc为底物,敲除lacZ和nanE阻断副反应途径减少底物浪费,过表达nagA、glmM和glmU以增强前体UDP-GlcNAc的供应,通过基因表达优化,3 L发酵罐中LNT Ⅱ产量为15.8 g/L,为利用生物质原料合成HMO提供了新思路。Sugita等[79]过表达大肠杆菌内源转运体基因setA显著提高了LNT Ⅱ产量,但SetA转运LNT Ⅱ具体转运机制尚不清晰。除了改造传统大肠杆菌,还可以改造益生菌大肠杆菌Nissle 1917合成LNT Ⅱ[80]。最近,Li等[81]首先确定了脑膜炎奈瑟氏菌MC58来源的LgtA具有合成LNT Ⅱ优势,基于高通量筛选平台确定高效突变体R13H/L24M/R205C,并在其C端融合InfB促融标签减少了包涵体形成,最后通过RBS工程和优化关键限速酶表达提高了LNT Ⅱ合成水平,在5 L发酵罐中产量达57.44 g/L,为目前报道最高水平。Wang等[82]采用两步法全细胞催化策略,开发了大肠杆菌-酵母耦合系统,以GlcNAc和乳糖为底物在5 L发酵罐中合成了52.34 g/L LNT Ⅱ,乳糖转化率达95.95%。
Other Images/Table from this Article
|