合成生物学 ›› 2023, Vol. 4 ›› Issue (1): 102-140.DOI: 10.12211/2096-8280.2022-030
光遗传学照进生物医学研究进展
于袁欢, 周阳, 王欣怡, 孔德强, 叶海峰
- 华东师范大学生命科学学院,上海市调控生物学重点实验室,华东师范大学医学合成生物学研究中心,上海 200241
-
收稿日期:2022-05-26修回日期:2022-09-05出版日期:2023-02-28发布日期:2023-03-07 -
通讯作者:叶海峰 -
作者简介:于袁欢(1992—),女,博士。研究方向为合成生物学与生物医学工程。
周阳(1994—),男,博士研究生。研究方向为合成生物学与生物医学工程。于袁欢 (1992—),女,博士。研究方向为合成生物学与生物医学工程。E-mail:yuyuanhuan@admin.ecnu.edu.cn
周阳(1994—),男,博士研究生。研究方向为合成生物学与生物医学工程。周阳 (1994—),男,博士研究生。研究方向为合成生物学与生物医学工程。E-mail:52191300036@stu.ecnu.edu.cn叶海峰 (1981—),男,研究员,博士生导师。主要从事合成生物学与生物医学工程领域的研究。利用合成生物学的理念和方法对细胞进行遗传学改造和重编程,重新设计、构建智能基因网络调控系统用于疾病的精准治疗。研究内容包括:遗传控制系统设计构建、智能细胞药物设计构建、光遗传学工具开发、精准可控的肿瘤免疫治疗、药物工程菌设计改造等。E-mail:hfye@bio.ecnu.edu.cn -
基金资助:国家重点研发计划(2019YFA0904500);国家自然科学基金(31971346);上海市科委(22N31900300)
Advances in optogenetics for biomedical research
YU Yuanhuan, ZHOU Yang, WANG Xinyi, KONG Deqiang, YE Haifeng
- Synthetic Biology and Biomedical Engineering Laboratory,Biomedical Synthetic Biology Research Center,Shanghai Key Laboratory of Regulatory Biology,Institute of Biomedical Sciences and School of Life Sciences,East China Normal University,Shanghai 200241,China
-
Received:2022-05-26Revised:2022-09-05Online:2023-02-28Published:2023-03-07 -
Contact:YE Haifeng
摘要:
近年来,光遗传学技术因具有非侵入性、可逆性、时空特异性等优点被广泛应用于生物医学研究领域,为疾病治疗提供了新思路和新理念。光作为一种理想的基因表达诱导物,以前所未有的时空精度操控基因表达和细胞行为。随着光遗传学技术的深入研究,基于光遗传学的个性化精准治疗和临床转化成为可能。本文主要介绍了响应不同波长的光遗传学工具及其用于神经系统疾病、肿瘤、心血管疾病、糖尿病、肠道疾病等精准治疗和用于控制基因转录表达、基因编辑、基因重组以及细胞器运动等应用。同时也介绍了光遗传学技术与智能电子设备的有机结合及其在便携式生物电子药物、人工智能诊疗方面的最新研究进展。光遗传学的迅速发展极大地拓展了传统生物电子医学领域。光控系统的远程可控性、可逆性和无毒性为光遗传学在生物医学中的应用提供了坚实的基础。这些方法的成功将对未来实践中的精准医疗产生持久的影响。最后探讨了光遗传学工具存在的问题和在未来临床应用面临的挑战,并对其未来发展前景进行了展望。
中图分类号:
引用本文
于袁欢, 周阳, 王欣怡, 孔德强, 叶海峰. 光遗传学照进生物医学研究进展[J]. 合成生物学, 2023, 4(1): 102-140.
YU Yuanhuan, ZHOU Yang, WANG Xinyi, KONG Deqiang, YE Haifeng. Advances in optogenetics for biomedical research[J]. Synthetic Biology Journal, 2023, 4(1): 102-140.
| 光响应元件 | 发光基团 | 光响应机理 | 来源 | 活化与失活光照条件/nm | 结合与解离半衰期 | 典型应用 | 参考 文献 |
|---|---|---|---|---|---|---|---|
| UVR8 | — | 解聚 | Arabidopsis thaliana | 300/黑暗 | ms/h | 控制趋化因子释放 | [ |
| UVR8-COP1 | — | 异源二聚 | Arabidopsis thaliana | 300/黑暗 | ms/h | 转基因表达 | [ |
| PhoCI | — | 光裂解 | Clavularia | 380/— | min/— | 控制蛋白质的清除 | [ |
| ChR2 | Retinal | 离子通道 | Chlamydomonas reinhardtii | 450/— | 0.2 ms/15 ms | 帕金森综合征等神经退行性疾病 | [ |
| Melanopsin | Retinal | 离子通道 | Mammalian retina | 450/— | ms/s | 糖尿病血糖控制 | [ |
| LOV | FMN | 结构变异 | Avena sativa | 450/黑暗 | 10 s/50 s | 钙离子信号控制、基因重组 | [ |
| AsLOV-Zdk1 | FMN | 解聚 | Avena sativa | 450/黑暗 | s/30~50 s | Notch信号控制、血糖控制 | [ |
| cpLOV-Zdk2 | FMN | 解聚 | Avena sativa | 450/黑暗 | s/30~50 s | 转录调控、细胞死亡 | [ |
| VVD | FMN/FAD | 同源二聚 | Neurospora crassa | 450/黑暗 | s/h | 控制RNA功能和代谢、基因重组、细胞消融 | [ |
| CRY2 | FAD | 同源二聚 | Arabidopsis thaliana | 450/黑暗 | s/min | 细胞坏死 | [ |
| CRY2clust | FAD | 同源二聚 | Arabidopsis thaliana | 450/黑暗 | s/min | 钙信号控制 | [ |
| CRY2olig | FAD | 同源二聚 | Arabidopsis thaliana | 450/黑暗 | s/min | 细胞通讯、钙信号控制 | [ |
| CRY2-CIB1 | FAD | 异源二聚 | Arabidopsis thaliana | 450/黑暗 | 10 s/12 min | 表观遗传控制、转录调控、基因重组、RNA修饰 | [ |
| nMag-pMag | FMN | 异源二聚 | Neurospora crassa | 450/黑暗 | 1.5 s/6.8 s | 转录调控、基因重组 | [ |
| EL222 | FMN | 同源二聚 | Erythrobacter litoralis | 450/黑暗 | s/s | 转录调控细胞迁移、细胞死亡 | [ |
| TtCBD | AdoCbl, MetCbl or CNCbl | 解聚 | Thermus thermophilus | 545/黑暗 | — | 细胞迁移、转基因表达 | [ |
| Dronpa | — | 解聚 | Pectiniidae | 500/400 | s/s | 控制Raf-MEK-ERK信号 | [ |
| PhyB-PIF3/PIF6 | PCB | 异源二聚 | Arabidopsis thaliana | 660/740 | 1.3 s/4 s | T细胞激活、转基因表达、基因重组 | [ |
| BphS | BV | 催化产生c-di-GMP | Rhodobacter sphaeroides | 680~810/— | 15 s/90 s | 转基因表达、糖尿病血糖控制 | [ |
| BphP1-PpsR2 | BV | 异源二聚 | Rhodopseudomonas palustris | 760/640 | 30 s/15 min | 转基因表达 | [ |
| PhyA-FHY1/FHL | PCB | 异源二聚 | Arabidopsis thaliana | 660/740 | — | 转基因表达、转录调控 | [ |
表1 哺乳动物细胞中常用光敏蛋白的特点
Table 1 Characteristics of photosensitive proteins commonly used in mammalian cells
| 光响应元件 | 发光基团 | 光响应机理 | 来源 | 活化与失活光照条件/nm | 结合与解离半衰期 | 典型应用 | 参考 文献 |
|---|---|---|---|---|---|---|---|
| UVR8 | — | 解聚 | Arabidopsis thaliana | 300/黑暗 | ms/h | 控制趋化因子释放 | [ |
| UVR8-COP1 | — | 异源二聚 | Arabidopsis thaliana | 300/黑暗 | ms/h | 转基因表达 | [ |
| PhoCI | — | 光裂解 | Clavularia | 380/— | min/— | 控制蛋白质的清除 | [ |
| ChR2 | Retinal | 离子通道 | Chlamydomonas reinhardtii | 450/— | 0.2 ms/15 ms | 帕金森综合征等神经退行性疾病 | [ |
| Melanopsin | Retinal | 离子通道 | Mammalian retina | 450/— | ms/s | 糖尿病血糖控制 | [ |
| LOV | FMN | 结构变异 | Avena sativa | 450/黑暗 | 10 s/50 s | 钙离子信号控制、基因重组 | [ |
| AsLOV-Zdk1 | FMN | 解聚 | Avena sativa | 450/黑暗 | s/30~50 s | Notch信号控制、血糖控制 | [ |
| cpLOV-Zdk2 | FMN | 解聚 | Avena sativa | 450/黑暗 | s/30~50 s | 转录调控、细胞死亡 | [ |
| VVD | FMN/FAD | 同源二聚 | Neurospora crassa | 450/黑暗 | s/h | 控制RNA功能和代谢、基因重组、细胞消融 | [ |
| CRY2 | FAD | 同源二聚 | Arabidopsis thaliana | 450/黑暗 | s/min | 细胞坏死 | [ |
| CRY2clust | FAD | 同源二聚 | Arabidopsis thaliana | 450/黑暗 | s/min | 钙信号控制 | [ |
| CRY2olig | FAD | 同源二聚 | Arabidopsis thaliana | 450/黑暗 | s/min | 细胞通讯、钙信号控制 | [ |
| CRY2-CIB1 | FAD | 异源二聚 | Arabidopsis thaliana | 450/黑暗 | 10 s/12 min | 表观遗传控制、转录调控、基因重组、RNA修饰 | [ |
| nMag-pMag | FMN | 异源二聚 | Neurospora crassa | 450/黑暗 | 1.5 s/6.8 s | 转录调控、基因重组 | [ |
| EL222 | FMN | 同源二聚 | Erythrobacter litoralis | 450/黑暗 | s/s | 转录调控细胞迁移、细胞死亡 | [ |
| TtCBD | AdoCbl, MetCbl or CNCbl | 解聚 | Thermus thermophilus | 545/黑暗 | — | 细胞迁移、转基因表达 | [ |
| Dronpa | — | 解聚 | Pectiniidae | 500/400 | s/s | 控制Raf-MEK-ERK信号 | [ |
| PhyB-PIF3/PIF6 | PCB | 异源二聚 | Arabidopsis thaliana | 660/740 | 1.3 s/4 s | T细胞激活、转基因表达、基因重组 | [ |
| BphS | BV | 催化产生c-di-GMP | Rhodobacter sphaeroides | 680~810/— | 15 s/90 s | 转基因表达、糖尿病血糖控制 | [ |
| BphP1-PpsR2 | BV | 异源二聚 | Rhodopseudomonas palustris | 760/640 | 30 s/15 min | 转基因表达 | [ |
| PhyA-FHY1/FHL | PCB | 异源二聚 | Arabidopsis thaliana | 660/740 | — | 转基因表达、转录调控 | [ |
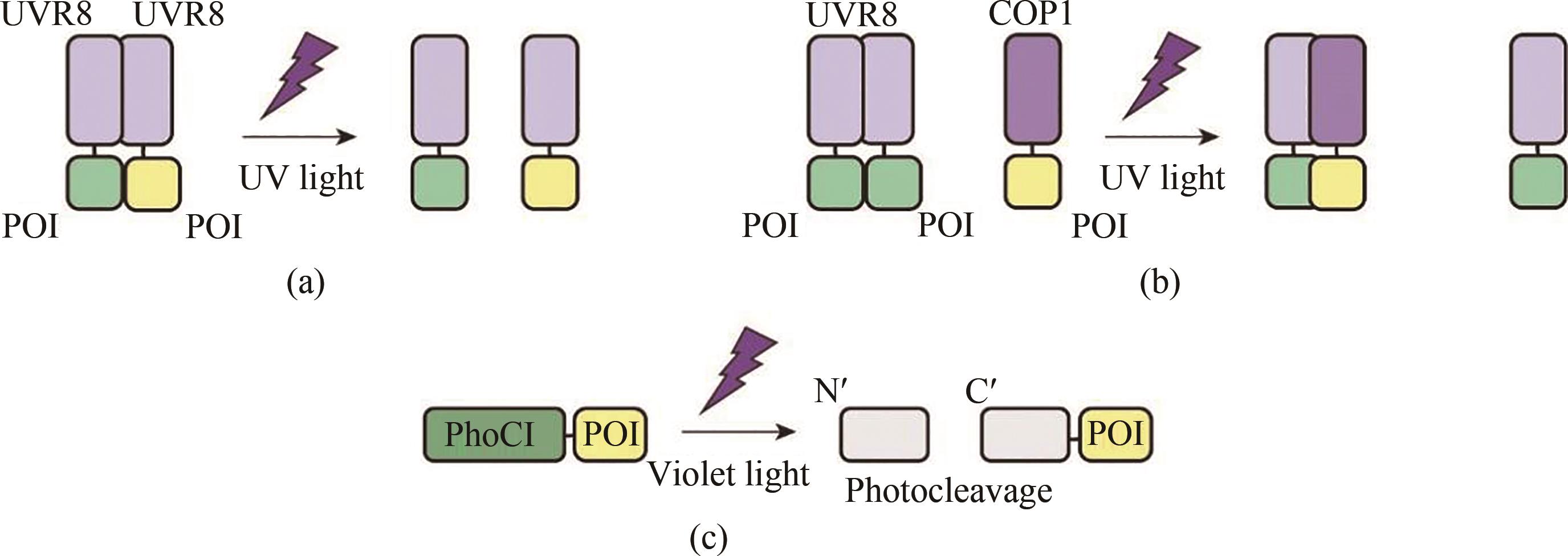
图1 紫外光和紫光响应的光遗传学工具工作原理示意图(a)紫外光刺激下,二聚化的UVR8分离为单体,与其相连的目的蛋白也随之解聚为单体。(b)黑暗条件下,二聚化的UVR8不能与其配体COP1结合;而在紫外光的刺激下,UVR8解聚,并与COP1蛋白结合,形成异源二聚体。(c)在紫光的照射下,荧光蛋白PhoCI发生断裂,与其相连的目的蛋白从而被释放,PhoCI也因此发生猝灭POI—目的蛋白
Fig. 1 Schematic diagram for the optogenetic tools responsive to ultraviolet and violet light(a) In the presence of UV light, homodimerized UVR8 dissociates into monomers and its fused proteins also depolymerizes. (b) Homodimerized UVR8 cannot bind to its ligand COP1 under dark conditions, but UVR8 binds to COP1 to form a heterodimer upon UV light illumination. (c) Violet light induces irreversible self-photocleavage of PhoC1 resulting in the quenching of green fluorescence.POI—protein of interest
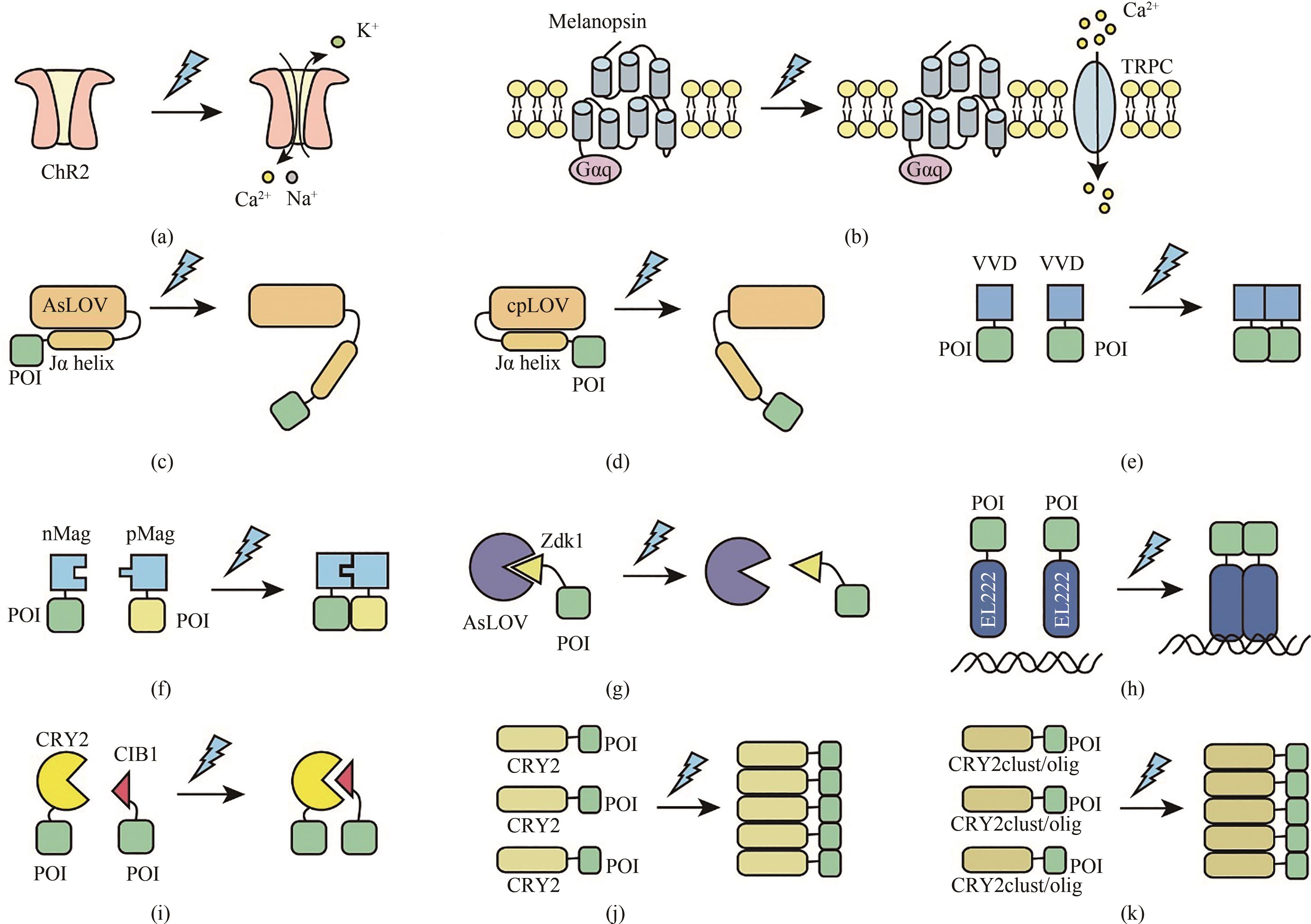
图2 蓝光响应的光遗传学工具工作原理示意图(a)蓝光响应的阳离子通道蛋白ChR2在蓝光刺激下激活,导致Ca2+、Na+等内流;(b)蓝光的照射下, 发色团视黄醛异质化, 与之相连的视黑素蛋白构象随之发生改变, 从而活化G蛋白(Gαq)、 磷脂酶C(PLC)和磷酸激酶C(PKC), 引起细胞膜上的瞬时受体电位离子通道(TRPCs)和内质网上的钙离子通道打开, 使钙离子快速内流;(c)在蓝光的刺激下,AsLOV 蛋白核心与黄素蛋白FMN之间的非共价作用转变为共价作用,产生光诱导的构象变化,解除了C端Jα 螺旋结构对LOV核心结构域的抑制作用;(d)在蓝光的刺激下,cpLOV 蛋白核心与黄素蛋白FMN之间的非共价作用转变为共价作用,产生光诱导的构象变化,解除了N端Jα 螺旋结构对LOV核心结构域的抑制作用;(e)蓝光刺激下,VVD蛋白发生同源二聚,与其相连蛋白的距离随之拉近;(f)在蓝光的刺激下,带正电的pMag蛋白与带负电的nMag蛋白发生异源二聚化,与其相连的目的蛋白也随之被拉近结合;(g)LOVTRAP是蓝光响应蛋白质分离系统。黑暗条件下,LOV2与Zdk1蛋白处于异源二聚化状态,蓝光照射下,LOV2蛋白结构发生改变,从而导致Zdk1蛋白与其分离;(h)在黑暗状态下, 光敏蛋白EL222 N端的LOV光感受域结合在其C端的HTH DNA结合域上, EL222不能二聚且无法与DNA结合,蓝光的刺激破坏了LOV和HTH的结合,EL222蛋白发生二聚并结合在靶DNA序列上;(i)在黑暗状态下,光敏蛋白CRY2与其配体蛋白CIB1处于分离状态,蓝光刺激下两者发生二聚;(j)在黑暗状态下,光敏蛋白CRY2处于分离状态,蓝光刺激下发生多聚;(k)在黑暗状态下,光敏蛋白CRY2clust/olig处于分离状态,蓝光刺激下发生多聚
Fig. 2 Schematic diagram for the optogenetic tools responsive to blue light(a) The blue light-responsive cation channel protein ChR2 is activated to induce influx of Ca2+ and Na+ by blue light illumination. (b) Under blue light illumination conditions, the chromophore retinal is isomerized to lead to the conformational changes of melanopsin thereby activating phospholipase C (PLC) through G protein (Gαq) and phospholipase C (PLC), which triggers Ca2+ influx by activating transient receptor potential ion channels (TRPCs) on the cell membrane and from the endoplasmic reticulum (ER). (c) With blue light illumination, a light-induced conformation is developed between the AsLOV protein core and the flavoprotein FMN, which results in undocking and unwinding of the LOV2 C-terminal Jα helix. (d) With blue light illumination, a light-induced conformation is formed between the cpLOV and the flavoprotein FMN, which leads to undocking and unwinding of the LOV2 C-terminal Jα helix. (e) Blue light induces the homodimerization of VVD, thus enabling proximity of the fused proteins. (f) Blue light induces the heterodimerization of pMag and nMag, thus enabling proximity of the fused target protein (g) LOVTRAP is a blue light responsive protein dissociation system, which is a reversible light-induced protein system. Zdk1 binds to LOV domain to form a heterodimer under dark conditions, but dissociates from LOV domain upon blue light illumination. (h) Under dark conditions, the N-terminal LOV photoreceptive domain of the light-sensitive protein EL222 binds to its C-terminal HTH DNA-binding domain, thus preventing EL222 dimerization and DNA binding, while blue light irradiation enables EL222 dimerization to recognize its target DNA sequences. (i) Blue light triggers heterodimerization of CRY2 and CIB1 to initiate gene expression. (j) Blue light triggers oligomerization of CRY2, while CRY2 forms a monomer under dark conditions. (k) Blue light triggers multimerization of CRY2clust/olig.
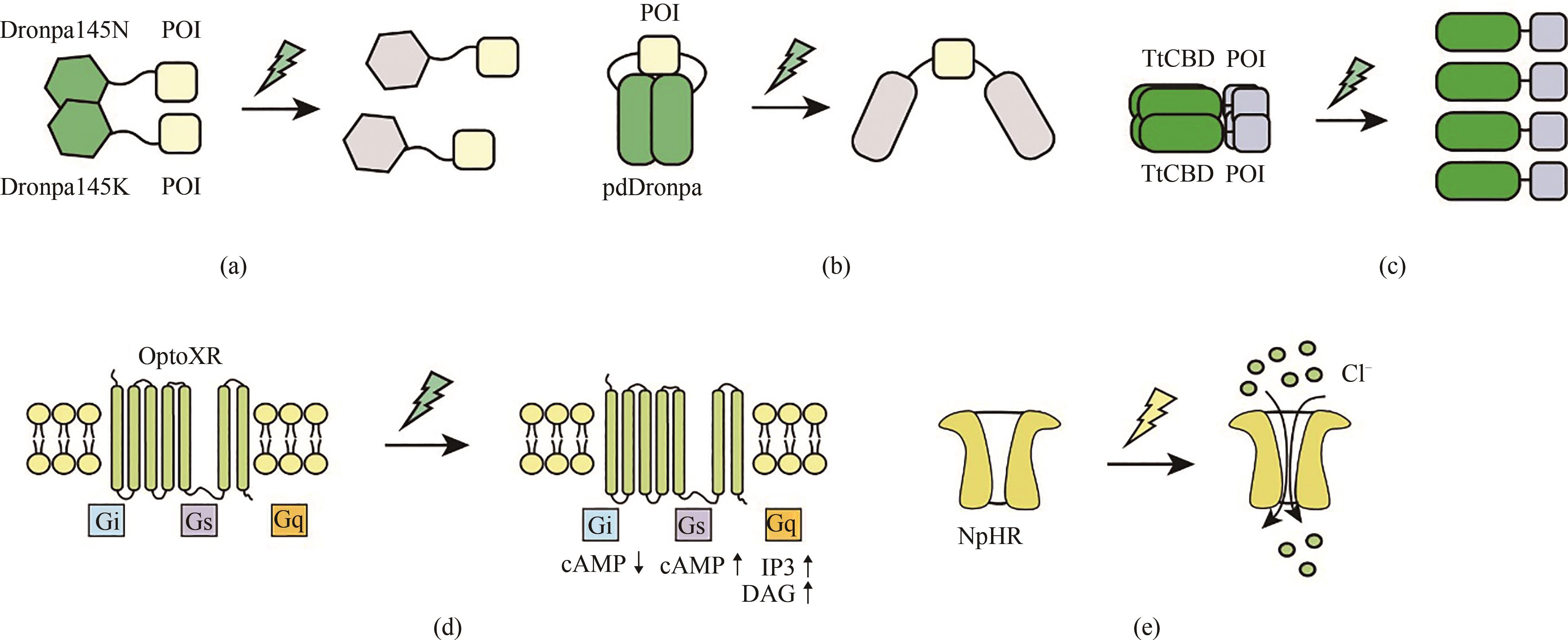
图3 青色光和黄色光响应的光遗传学工具工作原理示意图(a)在黑暗条件下,荧光蛋白Dronpa145N和Dronpa145K蛋白处于二聚化状态;青光照射下,Dronpa蛋白结构发生改变,呈现为非荧光状态,同时解聚为单体;(b)在黑暗条件下,光响应荧光蛋白pdDronpa处于同源二聚状态,与其相连目的蛋白的活性位点被阻挡,无法发挥生物学活性,青光照射使其解聚,从而使得目的蛋白活性位点暴露;(c)在黑暗条件下,TtCBD光敏蛋白处于同源四聚化状态,青光的照射使其解聚为单体;(d)嵌合视蛋白optoXRs在绿光(500 nm)刺激下发生构象变化,从而激活下游信号通路。(e)NpHR在黄色光刺激下被激活,引发Cl-内流
Fig. 3 Schematic diagram for the optogenetic tools responsive to cyan and yellow light(a) The fluorescent proteins Dronpa145N and Dronpa145K dimerizes under dark conditions, while the Dronpa protein undergoes a conformational change that enables depolymerization into monomers in the presence of cyan light. (b) Under dark conditions, pdDronpa homodimerizes and thus blocks the active sites of target proteins, while blue light illumination causes dissociation of the homodimers to expose the active sites of target proteins. (c) Cyan light illumination causes dissociation of the homotetramerized TtCBD. (d) Synthetic light-gated GPCRs (Opto-XRs) undergoes a conformational change that enables the activation of G-protein-mediated intracellular signaling cascades by cyan light illumination. (e) Yellow light-mediated activation of NpHR which triggers Cl- infux.

图4 红光响应的光遗传学工具工作原理示意图(a)光敏蛋白PhyB在黑暗条件下处于失活状态(Pr),红光作用导致其转变为激活状态(Pfr),Pfr状态下的PhyB蛋白在光敏色素PCB的帮助下,与其互作蛋白PIF结合为异源二聚体;(b)截短版的光敏蛋白ΔPhyA在黑暗条件下处于失活状态(Pr),红光作用导致其转变为激活状态(Pfr),Pfr状态下的PhyA蛋白在光敏色素PCB的帮助下,与其互作蛋白FHY1结合为异源二聚体;(c)黑暗状态下,BphP1以同源二聚体的形式存在;近红外光作用下,BphP1发生解聚并与PpsR2形成异源二聚体
Fig. 4 Schematic diagram for the optogenetic tools responsive to red/far red light(a) Phytochrome B (PhyB) maintains at Pr form which is biologically inactive in the dark. Red light illumination converts PhyB into the Pfr form and induces heterodimerization with PIF6 in the presence of the photosensitive pigment PCB. (b) In the dark, truncated Phytochrome A (ΔPhyA) maintains at Pr form, red light illumination converts ΔPhyA into the Pfr form and induces heterodimerization with FHY1 in the presence of the photosensitive pigment PCB. (c) Red light induces dissociation of homodimerized BphP1 which can interact with PpsR2 to form a heterodimerization pair.
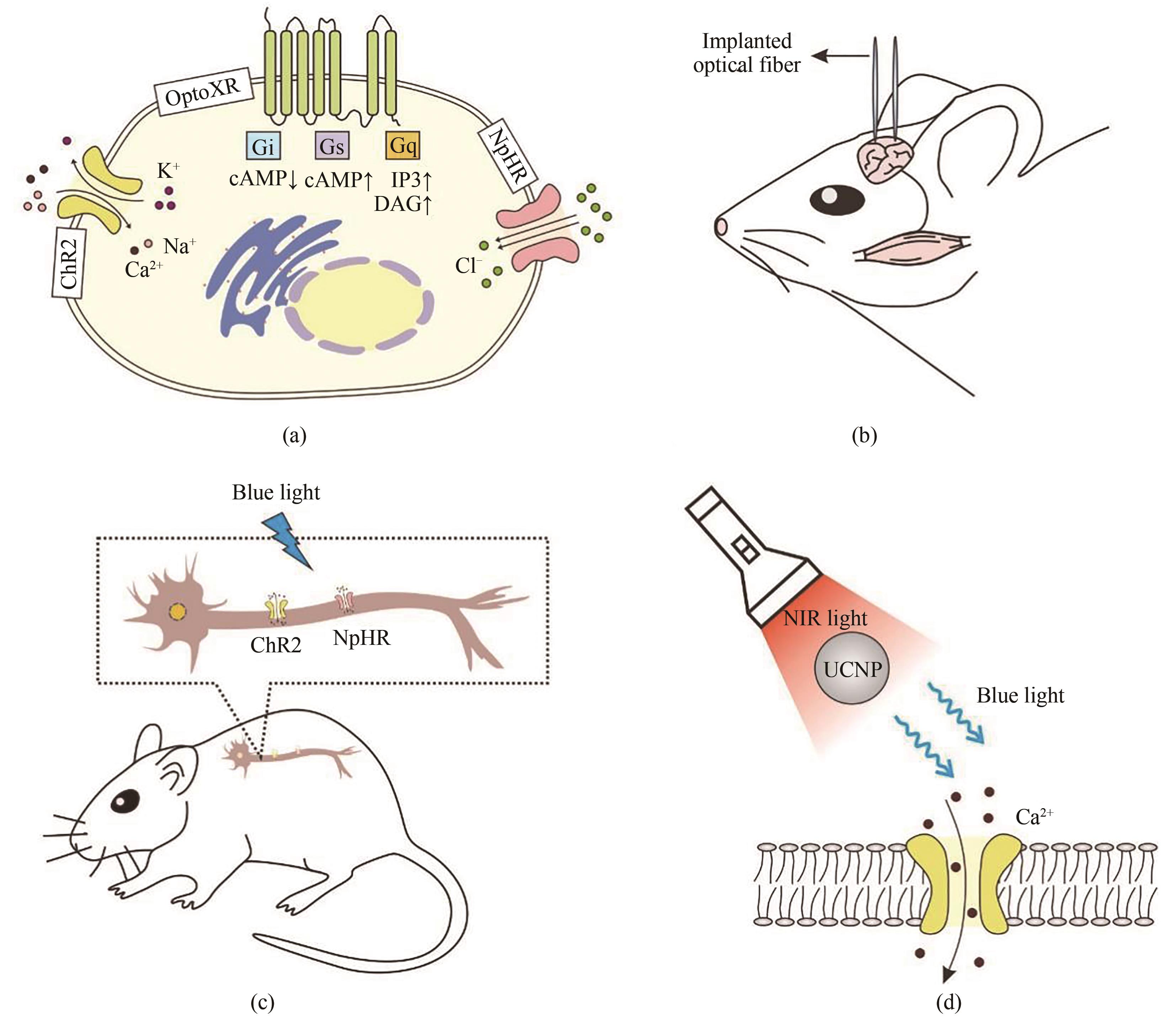
图5 光遗传学工具在神经生物学中的开发和应用(a)神经生物学领域中常用的三种光敏蛋白。ChR2蛋白在蓝光的刺激下可介导Ca2+、Na+等阳离子内流从而触发动作电位。NpHR蛋白可以在黄色光(580~590 nm)的刺激下介导Cl-内流,进而使得神经元超极化抑制神经元的活动。OptoXR蛋白是将视紫红质(rhodopsin)的胞内域替换成不同G蛋白偶联受体(GPCRs)的胞内域而形成。这些改造过的optoXRs可以在绿光(500 nm)的刺激下改变自身构象来激活下游的生化级联反应。(b)将ChR2基因导入小鼠大脑杏仁核区域里的两组神经中,通过植入光纤对其进行光学控制,发现小鼠猎杀行为神经通路。(c)将ChR2/NpHR基因导入小鼠背部的神经元中,通过光控该特定神经元的激活和抑制,发现小鼠瘙痒的相关神经通路。(d)通过AAV将ChR2基因导入到小鼠深部脑神经元中,并将UCNPs植入小鼠大脑深处。在体外使用近红外光照射时,UCNPs可以将近红外光转化成蓝光,进而激活大脑深处神经元上的ChR2蛋白,引发神经元活化
Fig. 5 Development and application of optogenetic tools in neurobiology(a) Three major types of opsins used in neurobiology. The blue light-responsive cation channel protein ChR2 is activated to induce influx of Ca2+ and Na+ and trigger an action potential. The yellow light-responsive NpHR is activated to allow chloride ions to enter the cytoplasm. Synthetic light-gated GPCRs (Opto-XRs) is engineered by replacing the intracellular domain of rhodopsin with that of G protein-coupled receptors to enable G-protein mediated intracellular signaling cascades. (b) Genetic targeting of ChR2 into the amygdala region of mouse brain enables optical control of the neural pathway of the mouse-hunting behavior by implanting optical fibers. (c) Genetic targeting of ChR2/NpHR into the cervical dorsal spinal cord of mice enables specific activation or suppression of the neurons, and avoids stimulation of non-targeted cells. (d) ChR2 is delivered into the deep brain neurons of mice by coupling with lanthanide-doped upconversion nanoparticles (UCNP), which can convert blue light to tissue-penetrable NIR light to activate ChR2 expressed dopaminergic neurons.
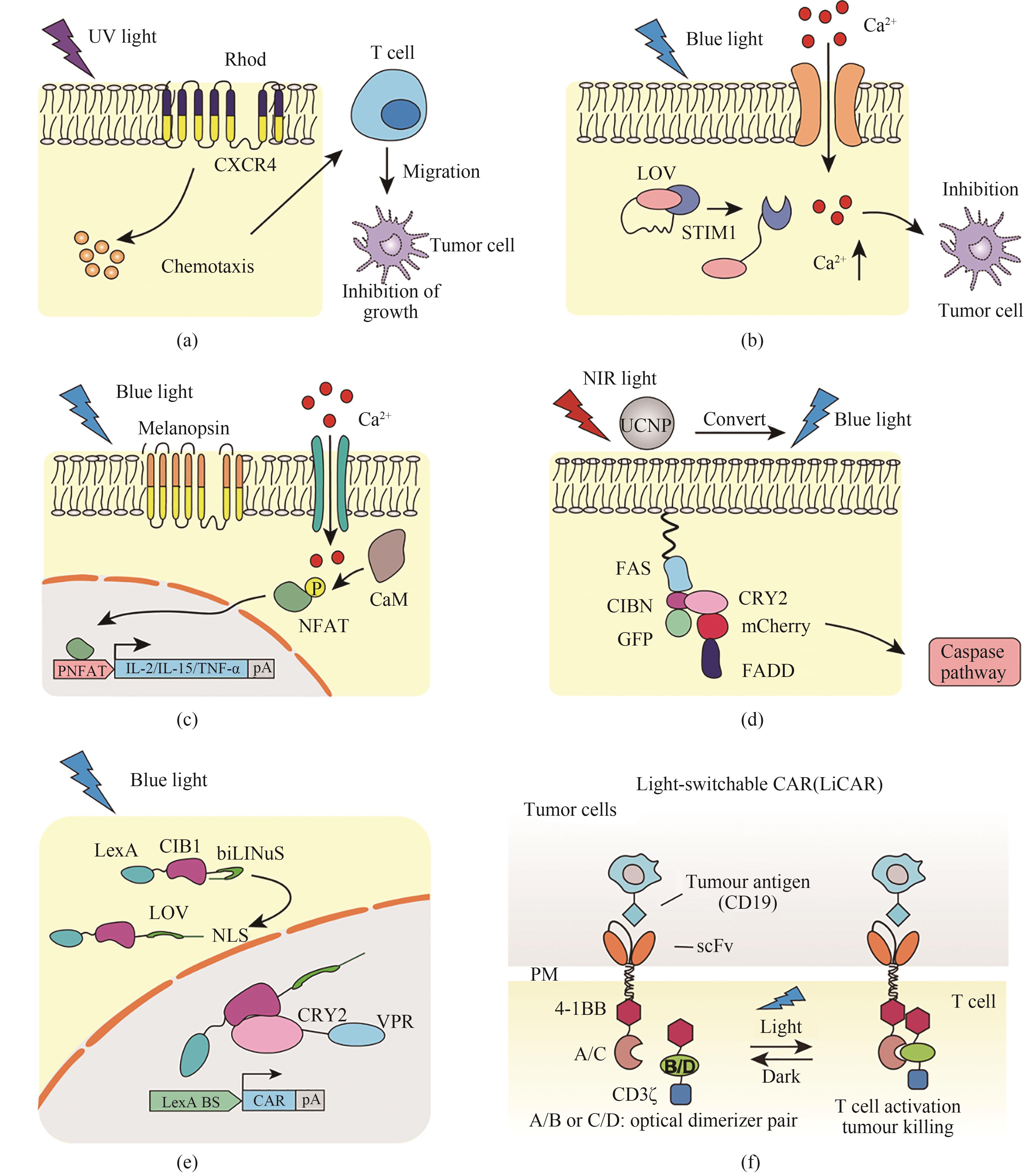
图6 光遗传学工具在肿瘤治疗中的应用(a)基于光控趋化因子受体的小鼠黑色素瘤免疫治疗。在505 nm波长的光照下,紫红质α亚基和趋化因子受体-4(CXCR4)组成的嵌合光激活趋化因子受体被激活,产生趋化因子信号并诱导T细胞极化,靶向至肿瘤部位,抑制肿瘤生长。(b)基于Opto-CRAC的细胞免疫治疗光遗传学系统。在蓝光的照射下,LOV2结构域羧基末端的Jα螺旋脱位,暴露出STIM1蛋白的C末端,从而刺激ORAI1 Ca2+通道,启动钙依赖级联反应,激活T细胞并诱导免疫应答,从而杀伤肿瘤。(c)基于视黑素蛋白的肿瘤免疫治疗光遗传学系统。在蓝光的照射下,视黑素蛋白促进钙离子内流,诱导钙依赖的NFAT信号通路启动,表达抗肿瘤因子IL-2/IL-15/TNF-α,从而抑制肿瘤生长。(d)光控细胞自噬系统用于肿瘤治疗。在上转化纳米材料UCNPs的作用下,组织穿透性更强的近红外光转化为蓝光,激活隐花色素系统CIBN与CRY2蛋白二聚化结合,从而将自噬相关蛋白FADD招募到质膜上的FAS蛋白,启动细胞自噬级联反应致使肿瘤细胞死亡。(e)LINTAD系统用于肿瘤治疗。蓝光照射后,LOV蛋白变构暴露出核定位信号NLS,从而将LexA-CIB1-biLINuS带入核内,与CIB1的光敏伴侣蛋白CRY2结合,共定位于报告系统的操纵序列上,激活CAR的表达。(f)LiCAR系统用于肿瘤治疗。光敏蛋白在蓝光照射下相互结合,从而招募CD3ζ结构域形成结构域完整的CAR受体,用于肿瘤细胞的识别和杀伤
Fig. 6 Optogenetic tools used for tumor therapy(a) Photochemokine receptors used for immunotherapy in murine melanoma. Under 505 nm light illumination, the chimeric photoactivated chemokine receptor composed of rhodopsin α subunit and chemokine receptor-4 (CXCR4) is activated to induce T cell polarization, resulting in the inhibition of tumor growth. (b) Opto-CRAC for cellular immunotherapy. Under blue light illumination, the Jα helix at the carboxyl terminus of LOV2 domain dislocates to expose the C-terminus of STIM1 protein, which stimulates the ORAI1 Ca2+ channel to initiate the calcium-dependent cascade and

图7 光遗传学工具在心血管疾病中的应用(a)蓝细菌光合系统用于治疗心肌缺血。将蓝细菌直接注射于急性心肌梗死模型大鼠心脏内。光照条件下,蓝细菌在光合作用中产生氧气,增加心肌细胞代谢活性,改善心室功能,从而缓解急性组织缺血;(b)光遗传学心脏起搏器。通过使用非选择性阳离子通道ChR2来控制心脏的兴奋性,从而实现光控心脏起搏;(c)抑制性光敏蛋白系统在心血管疾病中的应用。在黄光照射条件下,Arch-T促使H+外流,抑制心肌活性,缓解心律失常;(d)调节心肌细胞活性的光遗传起搏器。视黑素是一种光激活的G蛋白偶联受体,在470 nm蓝光照射下,能够激活磷脂酶C活性,催化PIP2水解产生IP3,导致Ca2+释放,增强心肌细胞的起搏活性
Fig. 7 Applications of optogenetic tools in treating cardiovascular diseases(a) Cyanobacterial photosynthetic systems for myocardial ischemia treatment. The cyanobacteria are injected into the hearts of the acute myocardial infarction model rats to produce oxygen through photosynthesis under light illumination conditions, which increases the metabolic activity of cardiomyocytes to improve ventricular function and alleviate acute tissue ischemia. (b) Optogenetic pacemakers. The non-selective cation channel ChR2 is used to control the cardiac excitability. (c) Inhibitory photosensitive protein systems for cardiovascular diseases treatment. ARCH-T mediated H+ efflux can inhibit myocardial activity and alleviate arrhythmia under yellow light illumination. (d) Optogenetic pacemakers for modulating cardiomyocyte activity. Melanopsin, a photoactivated G-protein-coupled receptor, activates the phospholipase C and catalyzes the hydrolysis of PIP2 to produce IP3, enabling release of Ca2+ and enhancement of the pacing activity of cardiomyocytes under blue light (470 nm) illumination conditions.
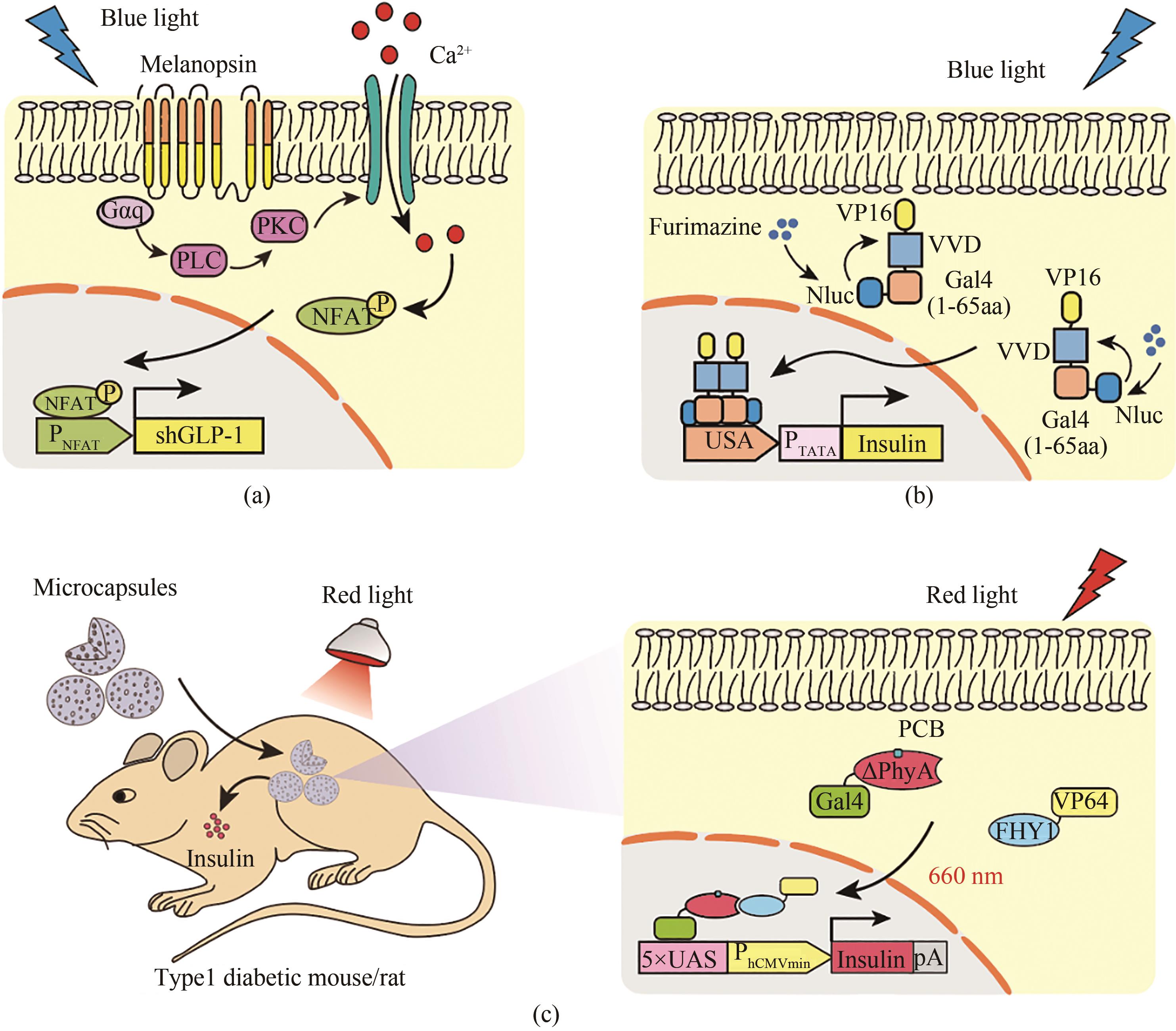
图8 治疗糖尿病的光遗传学工具(a)基于视黑素的蓝光调控系统对糖尿病的治疗。在蓝光的刺激下,视黑素(Melanopsin)构象发生改变,从而活化G蛋白(Gαq),磷脂酶C(PLC)和磷酸激酶C(PKC),导致瞬时受体电位离子通道(TRPCs)和胞内细胞器(内质网)钙离子通道打开,引起钙离子的快速内流,使NFAT磷酸化并入核,从而激活NFAT响应的启动子(PNFAT),启动GLP-1基因的表达,达到控制血糖稳态的效果。(b)基于光-氧-电压结构域的光受体蛋白对糖尿病的治疗。加入底物后,荧光素酶催化发出蓝光,光敏蛋白Vivid二聚化,与其相连的Gal4(1-65 aa)识别结构域随之发生二聚,入核并作用于其DNA结合位点5×UAS操纵序列上,与其融合的转录激活子VP16招募转录因子激活胰岛素的表达, 从而达到降血糖的效果。(c)基于ΔPhyA的红光调控系统对糖尿病的治疗。红光刺激下,ΔPhyA-GAL4与FHY1-VP64二聚结合形成复合体,入核并作用于其DNA结合位点5×UAS操纵序列上启动胰岛素的表达。将含有该系统的工程化细胞通过微胶囊包裹移植到糖尿病鼠的背部皮下,并给予光照,实现了红光介导的血糖稳态控制
Fig. 8 Optogenetic tools for diabetes therapy(a) Melanopsin-based blue light regulatory systems for diabetes treatment. Upon blue light stimulation, the melanopsin conformation is changed, thereby activating phospholipase C (PLC) through G protein (Gαq) and phospholipase C (PLC), which triggers Ca2+ influx by the activation of transient receptor potential ion channels (TRPCs) on the cell membrane and from the endoplasmic reticulum (ER). The activated NFAT translocates into the nucleus, and binds to its specific promoter (PNFAT), which initiates the GLP-1 gene expression to control blood glucose homeostasis. (b) The light-oxygen-voltage domain-based blue light regulatory system for diabetes treatment. In the presence of substrate, blue light is produced by the luciferase-catalyzed reaction, leading to the dimerization of the photosensitive protein Vivid. The DNA binding domain Gal4 (1-65 aa) fuses with Vivid and VP16 for incorporation into the nucleus to bind to the DNA operator (5 × UAS), which initiates the expression of insulin to control blood glucose homeostasis. (c) ΔPhyA-based red light regulatory systems for diabetes treatment. Under red light illumination, the hybrid transactivator FHY1-VP64 can be translocated into nucleus by photosensitive DNA binding elements (ΔPhyA-Gal4), in which it can bind to a particular operon sequence (5×UAS) to initiate the expression of insulin. The microcapsules containing engineered cells are implanted into the back of diabetic mice, through which the cells can be induced to produce insulin to control blood glucose homeostasis under red light illumination conditions.
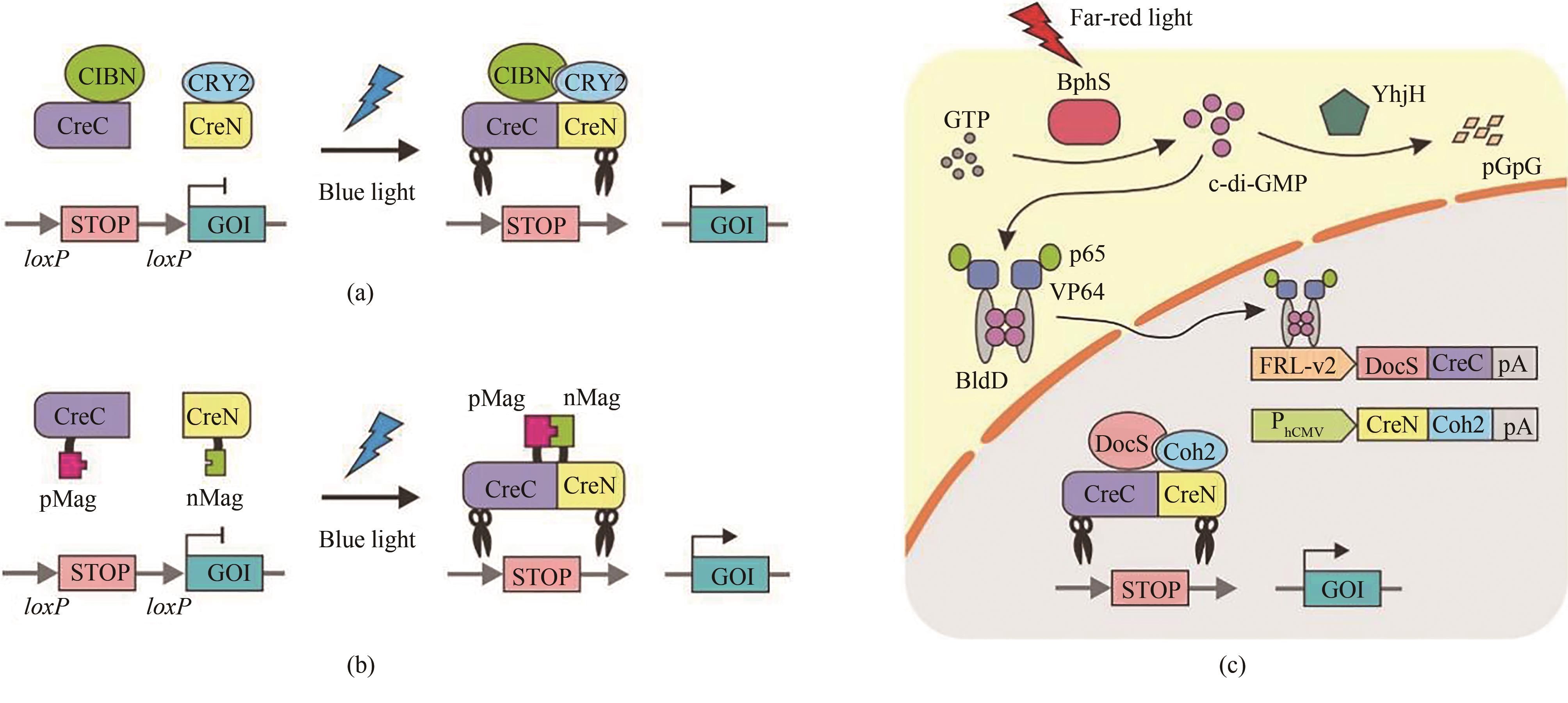
图9 基于split-Cre重组酶的光控基因重组系统(a)基于CRY2-CIBN的光控Cre重组酶系统。黑暗状态下,由于CreC和CreN被分成无活性的两部分,因而不具有基因重组功能。在蓝光照射下,CRY2发生构象改变与CIBN结合,拉动CreC和CreN形成完整有活性的Cre重组酶对相关基因进行重组。(b)基于pMag-nMag的光控Cre重组酶系统。在蓝光照射下,pMag与nMag结合,拉动CreC和CreN形成完整的Cre重组酶,进而对相关基因进行重组。(c)基于BphS-BldD的远红光调控基因重组系统(FISC系统)。在远红光的照射下,光敏蛋白BphS将细胞中的GTP转变成c-di-GMP,引发融合了转录激活子p65-VP64的BldD蛋白二聚化入核,激活DocS-CreC融合蛋白的表达。CreN-Coh2融合蛋白则由CMV启动子强启动表达。CreN和CreC在Coh2-DocS两个自发强相互作用蛋白的拉动下结合形成完整Cre重组酶,进而行使功能
Fig. 9 Light-controlled DNA recombination based on split-Cre recombinase(a) Photoactivatable Cre recombinase systems based on Cry2-CIBN. Under dark conditions, Cre recombinase is dissociated into two inactive parts. Under blue light illumination, CRY2 undergoes a conformational change and binds to CIBN, which induces CreC and CreN to form a complete Cre recombinase for restoration of its activity. (b) Photo-controlled Cre recombinase systems based on Magnet (pMag and nMag). Blue light illumination enables the dimerization of pMag and nMag to reconstitute a complete Cre recombinase to permit light-dependent DNA recombination. (c) A far-red light-inducible split Cre-loxP (FISC) system. Under far-red light illumination, the photosensitive protein BphS converts GTP into c-di-GMP within cells, which triggers the dimerization of BldD to fuse with transcriptional activator p65-VP64 for translocation into the nucleus to initiate the expression of DocS-CreC. Constitutive expressed CreN-Coh2 is driven by the CMV promoter. The catalytic activities of Cre recombinase can be restored through affinity interactions of their respective Coh2 and DocS fusion domains.

图10 基于CRISPR-Cas9的光控基因编辑系统(a)基于pMag-nMag的光控CRISPR-Cas9系统。黑暗状态下,NCas9和CCas9以无活性状态存在,不具有活性;而在蓝光照射下,pMag与nMag结合,拉动NCas9和CCas9形成完整的Cas9核酸酶对相关基因进行编辑。(b)基于Protected-sgRNA的光控CRISPR-Cas9系统。在黑暗情况下,sgRNA的种子序列被一个能被365 nm紫外光诱导切割的寡核苷酸链结合;在紫外光照射下,该寡核苷酸断裂,使sgRNA的种子序列暴露出来,从而引导Cas9切割相关DNA。(c)基于UCNPs材料的近红外光调控的CRISPR-Cas9系统。该系统将Cas9-sgRNA的复合物通过PEI,SiO2材料包裹在UCNPs材料外侧,UCNPs可以将980 nm近红外光转换为紫外光,紫外光可引发Cas9-sgRNA复合物的彻底释放,进而对相关基因进行编辑。(d)基于APC金纳米颗粒的第二近红外窗口光调控的CRISPR-Cas9系统。该APC金纳米颗粒可携带热诱导启动子启动的Cas9质粒,并将其高效运输至小鼠体内器官和组织。在1064 nm的第二近红外窗口光照射下,APC金纳米颗粒可以将光能转化为热能,激活其携带的热启动质粒表达Cas9核酸酶,从而发挥基因编辑功能。(e)基于BphS-BldD的远红光调控基因编辑系统
Fig. 10 Light-controlled gene editing systems based on CRISPR-Cas9(a) Photoactivatable CRISPR-Cas9 systems based on pMag-nMag. Under dark conditions, Cas9 is splitted into two fragments without nuclease activity. With blue light illumination, the NCas9 and CCas9 domains can be reassociated to form a complete Cas9 by the light-dependent dimerization of pMag and nMag, thereby reconstituting editing for targeted genes. (b) Photoactivatable CRISPR-Cas9 systems based on protected sgRNA. Under dark conditions, the seed sequence of sgRNA is bound by an oligonucleotide that could be cleaved by UV light. With UV light illumination, the oligonucleotide is broken, and the seed sequence of sgRNA is exposed to allow Cas9 to bind to and cleave the target DNA. (c) Near-infrared light-regulated CRISPR-Cas9 systems based on UCNPs. The Cas9-sgRNA complex is wrapped on the outside of UCNPs by PEI and SiO2, which have the capability to convert near-infrared light (980 nm) to ultraviolet light for the Cas9-sgRNA complex to bind to target genes. (d) CRISPR-Cas9 systems based on APC gold nanoparticles. Cas9 plasmids carrying the heat-induced promoter can be efficiently delivered into mice by APC gold nanoparticles. Under near infrared light irradiation at 1064 nm, APC gold nanoparticles can convert light energy to heat, which activates the expression of Cas9 nuclease for genome editing. (e) A far-red light-activated split-Cas9 (FAST) system. The far-red light activates the expression of the fusion proteinNCas9-Coh2, and the CMV promoter drives the expression of the fusion protein DocS-CCas9, which consequently activates Cas9 nuclease by heterodimerization between Coh2 and DocS.

图11 基于CRISPR-dCas9的光控基因转录系统(a)基于CRY2-CIBN的光控dCas9基因转录系统。黑暗情况下,N端dCas9和C端dCas9处于无活性的分离状态;当蓝光照射时,CRY2发生构象改变与CIBN结合,带动融合在CRY2蛋白上的转录激活子VP64激活下游基因的表达。(b)基于SAM和pMag-nMag的蓝光调控dCas9基因转录系统。SAM是指在sgRNA的茎环结构上插入了能被MS2蛋白识别的MS2-box序列,使得融合了p65-HSF1复合型转录激活子的MS2蛋白可以发挥转录激活的功能。在蓝光的照射下,由pMag和nMag蛋白的相互作用拉动N端dCas9和C端dCas9结合形成完整有活性的dCas9,并在SAM系统的作用下就激活相关基因转录表达。(c)基于SAM和BphS-BldD的远红光调控基因转录系统(FACE系统)。黑暗状态下,无法激活基因转录;只有在远红光的照射下,才可诱导SAM系统的MS2-p65-HSF1融合蛋白表达,进而通过sgRNA-dCas9系统的定位作用激活相关基因的表达。(d)基于REDMAP的光控dCas9基因转录系统。黑暗状态下,无法激活基因的转录;在红光的照射下,△PhyA与FHY1形成异源二聚体,启动SAM系统的MS2-p65-HSF1融合蛋白的表达;进而通过sgRNA-dCas9系统的定位作用实现对靶基因的转录激活(FAST)。原理同图9(c),远红光激活NCas9-Coh2融合蛋白的表达,CMV启动子强启动表达DocS-CCas9融合蛋白。NCas9和CCas9在Coh2-DocS两个自发强相互作用蛋白的拉动下结合形成完整的Cas9核酸酶,进而行使基因编辑功能
Fig. 11 Light-controlled gene transcription systems based on CRISPR-dCas9(a) Photoactivable dCas9-meidated transcription systems based on CRY2-CIBN. Under dark conditions, dCas9 is splitted into two fragments without catalytic activity. With blue-light illumination, CRY2 undergoes a conformational change that enables interactions with CIBN, which causes translocation of the transactivator VP64 to activate downstream gene transcription. (b) Blue light-controlled dCas9-meidated gene transcription systems based on SAM and pMag-nMag. SAM, a synergistic activation mediator that extends guide RNAs with an insertion of a MS2-box sequence into the loop of gRNA, can recruit effector protein to initiate gene transcription. Under blue-light illumination conditions, the NdCas9 and CdCas9 domains can be reassociated to form a complete dCas9 by the light-dependent dimerization of pMag and nMag, thereby activating the downstream gene transcription. (c) A far red light-controlled gene transcription system based on SAM and BphS-BldD. Under far-red light illumination conditions, the fused protein MS2-p65-HSF1 can express to activate the target gene transcription. (d) Light-controlled dCas9-midated gene transcription systems based on REDMAP. Under red-light illumination conditions, the heterodimerization of ΔPhyA and FHY1 enables the expression of the fused protein MS2-p65-HSF1 to activate endogenous gene expression by sgRNAs-mediated recruitment of the transcriptional activator domain.

图12 基于CRISPR-Cas12a/dCas12a技术的光控基因转录和基因编辑系统(a)蓝光诱导的分割型Cas12a基因编辑系统。在蓝光照射下,光敏蛋白pMag与nMagHigh1形成二聚体,拉动N端LbCas12a和C端LbCas12a结合形成完整的Cas12a,使其恢复核酸酶活性,并在crRNA的引导下实现对目的DNA的切割。(b)远红光调控的Cas12a基因编辑系统。光敏蛋白BphS在远红光照射下将GTP转化为c-di-GMP,使得人工合成的转录激活子p65-HSF1-BldD同源二聚,并转位到细胞核内与操作子结合,诱导 Cas12a表达,并在crRNA的引导下切割目的DNA。(c)基于SunTag和BphS-BldD的远红光调控的dCas12a基因转录系统。同样地,SunTag系统中的dCas12a- GCN4只有在远红光照射时才可以表达,通过招募scFv融合表达的激活子,在crRNA的引导下靶向目的基因位点,实现对靶基因的激活
Fig. 12 Light-controlled gene transcription and editing systems based on CRISPR-Cas12a/dCas12a(a) Blue light-controlled split-Cas12a gene editing systems. Under blue-light illumination conditions, pMag and nMagHigh1 that are fused to dCas12a are reassociated by the dimerization of pMag and nMag, thereby recovering the catalytic activity of Cas12a to cleave the target DNA sequence. (b) Far-red light-controlled Cas12a gene editing systems. Under far-red light illumination conditions, BphS can convert GTP into c-di-GMP to trigger the dimerization of p65-HSF1-BldD for binding with the operator to induce the expression of Cas12a for targeted genome cleavage. (c) Far-red light-controlled gene transcription systems based on SunTag and BphS-BldD. Under far-red light illumination conditions, the expression of the fusion protein dCas12a-GCN4 can be induced to activate the target gene transcription by the recruitment of the transactivator fused with ScFv.
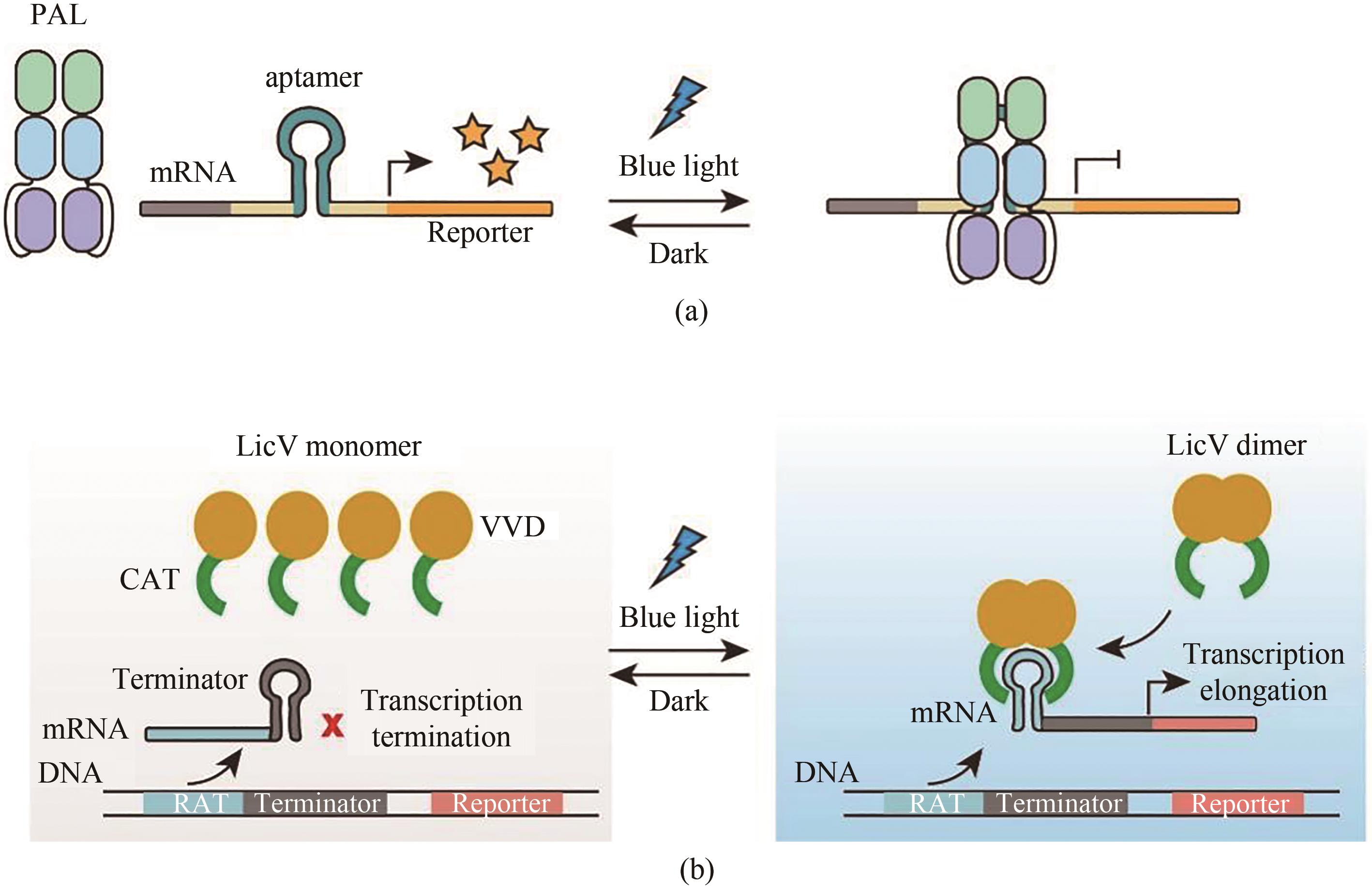
图13 光控RNA水平的基因表达调控系统(a)基于PAL蛋白的转录调控系统。黑暗状态下,PAL蛋白与RNA适体分离,报告基因表达;在蓝光照射下,PAL蛋白与RNA适体结合,阻碍报告基因的表达;(b)基于LicV蛋白的转录调控系统。mRNA上存在一段茎环结构的终止序列,阻碍了报告基因的表达,当蓝光照射时,VVD蛋白二聚,导致CAT形成二聚体,并与核糖核酸抗终止子序列(RAT)结合,打开了茎环结构,从而恢复了报告基因的表达
Fig. 13 Light-controlled transgene expression systems based on RNA level(a) Light-controlled transgene expression systems based on PAL. Under dark conditions, PAL is dissociated from the RNA aptamer to initiate reporter gene expression. With blue light illumination, PAL can bind to RNA aptamer, which can suppress the reporter gene expression. (b) Light-controlled transgene expression systems based on LicV. A stem-loop inserted termination sequence hinders the reporter gene expression, and under blue light illumination, the dimerized VVDs enables dimerization of CATs, which binds to the RAT sequence and unfold the stem-loop, thereby restoring the reporter gene expression.
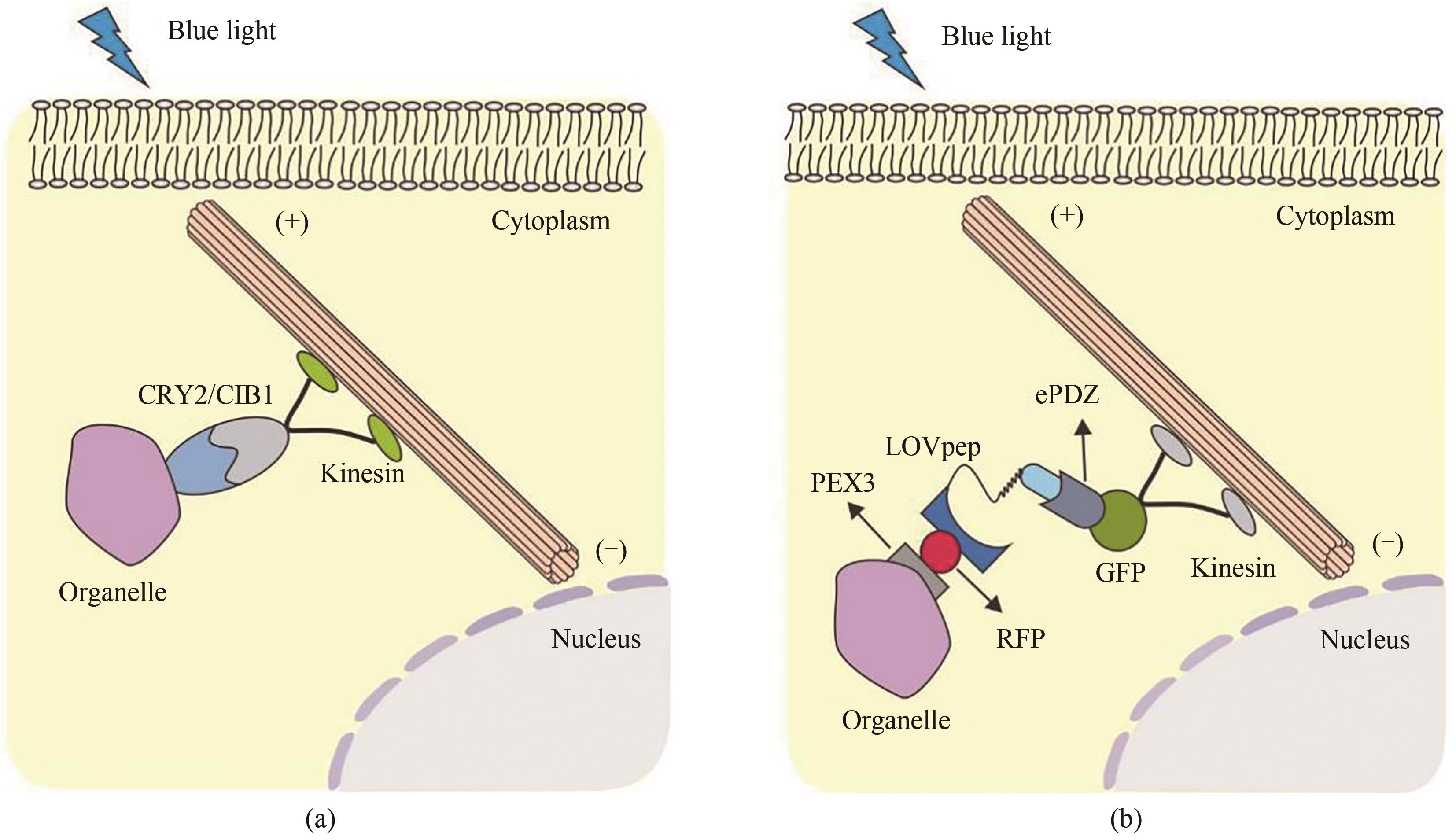
图14 光控细胞器运动和定位(a)基于CRY2-CIB1的光控细胞器定位系统。黑暗情况下,CRY2和CIB1蛋白处于游离状态,细胞器和分子马达没有相互作用;当接受到蓝光刺激后,融合有细胞器的CRY2蛋白和融合有马达分子的CIB1蛋白发生异源二聚,从而使得细胞器在分子马达的作用下被运送到细胞核或者细胞膜。(b)基于LOV的光控细胞器定位系统。黑暗情况下,LOV蛋白处于游离状态;在蓝光的照射下,融合有PEX-LOV的细胞器会与工程化PDZ结构域ePDZb1(融合了分子马达)结合, 在分子马达的驱动下, 实现细胞器的转运或再定位
Fig. 14 Light-controlled movement and localization of organelles(a) Light-controlled organelle localization based on CRY2-CIB1. Under blue-light illumination conditions, CRY2 undergoes a conformational change that enables interactions with CIBN for the movement of organelles driven by the molecular motor. (b) Light-controlled localization of organelles based on LOV. Under blue light irradiation conditions, the organelles fused with PEX-LOV can bind to the engineered PDZ domain ePDZb1fused with molecular motor, leading to the movement and localization of organelle.

图15 光遗传学工具用于智能化生物电子医疗(a)大脑控制转基因表达的无线供电光遗传学植入系统。该大脑思维控制的转基因表达装置可以通过人类大脑活动无线控制细胞中的基因表达。脑电图(EEG)耳机捕捉脑电波活动,并通过蓝牙传输到场强发生器接口(BCI),调节近红外发光二极管的开启,从而控制近红外光激活细菌双胍酸环化酶(DGC),将GTP转化为c-di-GMP,激活干扰素基因刺激因子(STING)依赖的信号通路,从而启动目的基因的表达。(b)基于光控定制化细胞的半自动糖尿病智能诊疗系统。血糖仪测得血糖数据后,可通过蓝牙将数据自动传输到智能控制器和智能手机上。智能控制器会根据预设的血糖值和光照强度针对血糖浓度的高低输出不同强度的远红光。在远红光照射下,细菌光感受器BphS将细胞内的GTP转化为c-di-GMP,使杂交型转录激活子p65-VP64-BldD二聚化入核上,结合到嵌合启动子上,启动胰岛素或胰高血糖素样肽基因的转录表达,从而控制血糖稳态。(c)可穿戴式智能电子手表控制的血糖稳态调控系统。该系统利用智能手表发射的绿光激活小鼠皮下的人工定制化细胞。手表背部发光二极管开启时,锚定在细胞膜上绿光响应的TtCBD蛋白解聚,TtCBD与杂交型转录激活子TetR-VPR从细胞膜掉落,入核并启动胰高血糖素样肽基因的转录表达,从而控制血糖稳态简单地说,脑电波(EEG)接收器捕捉到特定的大脑电波,并将信号传递给脑-计算机接口(BCI),当接收到的脑电波超过一定阈值时,BCI会做出响应,打开近红外LED灯,包含光响应基因环路的细胞则会启动下游报告基因的表达,产生报告蛋白分泌型碱性磷酸酶(SEAP)。因此,通过脑机接口技术和光遗传学技术结合,可以实现通过人类的意念控制基因的表达。该光响应基因环路主要由一种光依赖性细菌二鸟苷酸环化酶(DGCL)组成,它能将细胞中的GTP转化为环二鸟苷酸单磷酸(c-di-GMP)[201]。c-di-GMP结合并激活内质网上的干扰素基因刺激因子(STING)[202],这进一步激活了TBK1介导的转激活因子IRF3的磷酸化。磷酸化的IRF3进入细胞核,与人工合成的启动子结合,驱动下游转基因表达。该研究工作概念性验证了通过意念控制报告基因的表达。
Fig. 15 Optogenetic tools for intelligent bioelectronic medicine(a) A brain-controlled wireless-powered optogenetic implant device for transgene expression. This brain-controlled transgenic expression device can wirelessly control gene expression through human brain activities. Electroencephalogram (EEG) headset captures brain wave activities and transmits them to the field intensity generator interface (BCI) through Bluetooth, which integrates with a near-infrared LED. The bacterial diguanylate cyclase (DGC) can be activated by near-infrared light, which converts GTP into c-di-GMP for the activation of the STING signal pathway to initiate the transgene expression. (b) Semi-automatic intelligent diagnosis and treatment systems based on optogenetic designer cells for diabetes treatment. The blood glucose value detected from the blood glucose monitor can be automatically transmitted to the smart controller and smart phone through Bluetooth. The smart controller can regulate far-red light intensity based on the blood glucose value. Under far-red light irradiation, BphS converts intracellular GTP into c-di-GMP, which dimerizes the hybrid transcriptional activator p65-VP64-BldD into the nucleus for binding to the chimeric promoter to initiate the expression of insulin or GLP-1 for controlling blood glucose homeostasis. (c) Wearable smart watch-controlled optogenetic systems for disbetes treatment. The system utilizes green light from a smartwatch to activate artificially customized cells implanted into the skin of mice. The green light-responsive TtCBD is anchored onto the cell membrane. When the LEDs are turned on, TtCBD is depolymerized, and the hybrid transcriptional activator TetR-VPR is separated from the cell membrane to initiate the transcription expression of GLP-1for controlling blood glucose homeostasis.
| 1 | BOYDEN E S, ZHANG F, BAMBERG E, et al. Millisecond-timescale, genetically targeted optical control of neural activity[J]. Nature Neuroscience, 2005, 8(9): 1263-1268. |
| 2 | BI A D, CUI J J, MA Y P, et al. Ectopic expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration[J]. Neuron, 2006, 50(1): 23-33. |
| 3 | NEWS STAFF THE. Insights of the decade. Stepping away from the trees for a look at the forest. Introduction[J]. Science, 2010, 330(6011): 1612-1613. |
| 4 | Method of the year 2010[J]. Nature Methods, 2011, 8(1): 1. |
| 5 | FAVORY J J, STEC A, GRUBER H, et al. Interaction of COP1 and UVR8 regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis [J]. The EMBO Journal, 2009, 28(5): 591-601. |
| 6 | WU Y I, FREY D, LUNGU O I, et al. A genetically encoded photoactivatable Rac controls the motility of living cells[J]. Nature, 2009, 461(7260): 104-108. |
| 7 | YE H F, DAOUD-EL BABA M, PENG R W, et al. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice[J]. Science, 2011, 332(6037): 1565-1568. |
| 8 | WANG X, CHEN X J, YANG Y. Spatiotemporal control of gene expression by a light-switchable transgene system[J]. Nature Methods, 2012, 9(3): 266-269 |
| 9 | KAINRATH S, STADLER M, REICHHART E, et al. Green-light-induced inactivation of receptor signaling using cobalamin-binding domains[J]. Angewandte Chemie International Edition, 2017, 56(16): 4608-4611. |
| 10 | LEVSKAYA A, WEINER O D, LIM W A, et al. Spatiotemporal control of cell signalling using a light-switchable protein interaction[J]. Nature, 2009, 461(7266): 997-1001. |
| 11 | HEYES D J, KHARA B, SAKUMA M, et al. Ultrafast red light activation of Synechocystis phytochrome Cph1 triggers major structural change to form the Pfr signalling-competent state[J]. PLoS One, 2012, 7(12): e52418. |
| 12 | KABERNIUK A A, SHEMETOV A A, VERKHUSHA V V. A bacterial phytochrome-based optogenetic system controllable with near-infrared light[J]. Nature Methods, 2016, 13(7): 591-597. |
| 13 | SHAO J W, XUE S, YU G L, et al. Smartphone-controlled optogenetically engineered cells enable semiautomatic glucose homeostasis in diabetic mice[J]. Science Translational Medicine, 2017, 9(387): eaal2298. |
| 14 | SHAO J W, WANG M Y, YU G L, et al. Synthetic far-red light-mediated CRISPR-dCas9 device for inducing functional neuronal differentiation[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(29): E6722-E6730. |
| 15 | CHEN X H, CHEN Y X, XIN H H, et al. Near-infrared optogenetic engineering of photothermal nanoCRISPR for programmable genome editing[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(5): 2395-2405. |
| 16 | NIHONGAKI Y, FURUHATA Y, OTABE T, et al. CRISPR-Cas9-based photoactivatable transcription systems to induce neuronal differentiation [J]. Nature methods, 2017, 14(10): 963-966. |
| 17 | POLSTEIN L R, GERSBACH C A. A light-inducible CRISPR-Cas9 system for control of endogenous gene activation[J]. Nature Chemical Biology, 2015, 11(3): 198-200. |
| 18 | YU Y H, WU X, GUAN N Z, et al. Engineering a far-red light-activated split-Cas9 system for remote-controlled genome editing of internal organs and tumors[J]. Science Advances, 2020, 6(28): eabb1777 |
| 19 | NIHONGAKI Y, KAWANO F, NAKAJIMA T, et al. Photoactivatable CRISPR-Cas9 for optogenetic genome editing[J]. Nature Biotechnology, 2015, 33(7): 755-760. |
| 20 | HEMPHILL J, BORCHARDT E K, BROWN K, et al. Optical control of CRISPR/Cas9 gene editing[J]. Journal of the American Chemical Society, 2015, 137(17): 5642-5645. |
| 21 | JAIN P K, RAMANAN V, SCHEPERS A G, et al. Development of light-activated CRISPR using guide RNAs with photocleavable protectors[J]. Angewandte Chemie International Edition, 2016, 55(40): 12440-12444. |
| 22 | BUGAJ L J, CHOKSI A T, MESUDA C K, et al. Optogenetic protein clustering and signaling activation in mammalian cells[J]. Nature Methods, 2013, 10(3): 249-252. |
| 23 | WAGNER J C, PLATT R J, GOLDFLESS S J, et al. Efficient CRISPR-Cas9-mediated genome editing in Plasmodium falciparum [J]. Nature Methods, 2014, 11(9): 915-918. |
| 24 | TAN P, HE L, HUANG Y, et al. Optophysiology: illuminating cell physiology with optogenetics[J]. Physiological Reviews, 2022, 102(3): 1263-1325. |
| 25 | GOVORUNOVA E G, SINESHCHEKOV O A, JANZ R, et al. Natural light-gated anion channels: a family of microbial rhodopsins for advanced optogenetics[J]. Science, 2015, 349(6248): 647-650. |
| 26 | HANKINS M W, PEIRSON S N, FOSTER R G. Melanopsin: an exciting photopigment[J]. Trends in Neurosciences, 2008, 31(1): 27-36. |
| 27 | WEISSENBERGER S, SCHULTHEIS C, LIEWALD J F, et al. PACα-an optogenetic tool for in vivo manipulation of cellular cAMP levels, neurotransmitter release, and behavior in Caenorhabditis elegans [J]. Journal of Neurochemistry, 2011, 116(4): 616-625. |
| 28 | NOGLY P, STANDFUSS J. Light-driven Na+ pumps as next-generation inhibitory optogenetic tools[J]. Nature Structural & Molecular Biology, 2015, 22(5): 351-353. |
| 29 | GLANTZ S T, BERLEW E E, JABER Z, et al. Directly light-regulated binding of RGS-LOV photoreceptors to anionic membrane phospholipids[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(33): E7720-E7727. |
| 30 | HUANG Z L, WU Y Q, ALLEN M E, et al. Engineering light-controllable CAR T cells for cancer immunotherapy[J]. Science Advances, 2020, 6(8): eaay9209. |
| 31 | ALI A M, REIS J M, XIA Y, et al. Optogenetic inhibitor of the transcription factor CREB[J]. Chemistry & Biology, 2015, 22(11): 1531-1539. |
| 32 | RIZZINI L, FAVORY J J, CLOIX C, et al. Perception of UV-B by the Arabidopsis UVR8 protein[J]. Science, 2011, 332(6025): 103-106. |
| 33 | KIM C K, CHO K F, KIM M W, et al. Luciferase-LOV BRET enables versatile and specific transcriptional readout of cellular protein-protein interactions[J]. eLife, 2019, 8: e43826. |
| 34 | PATEL A L, YEUNG E, MCGUIRE S E, et al. Optimizing photoswitchable MEK[J]. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(51): 25756-25763. |
| 35 | JUNG H, KIM S W, KIM M, et al. Noninvasive optical activation of Flp recombinase for genetic manipulation in deep mouse brain regions[J]. Nature Communications, 2019, 10: 314. |
| 36 | YAO S Q, YUAN P, OUELLETTE B, et al. RecV recombinase system for in vivo targeted optogenomic modifications of single cells or cell populations[J]. Nature Methods, 2020, 17(4): 422-429. |
| 37 | ENDO M, IWAWAKI T, YOSHIMURA H, et al. Photocleavable cadherin inhibits cell-to-cell mechanotransduction by light[J]. ACS Chemical Biology, 2019, 14(10): 2206-2214. |
| 38 | ZHANG X L, DONG C M, HUANG W Y, et al. Rational design of a photo-responsive UVR8-derived protein and a self-assembling peptide-protein conjugate for responsive hydrogel formation[J]. Nanoscale, 2015, 7(40): 16666-16670. |
| 39 | CHEN D, GIBSON E S, KENNEDY M J. A light-triggered protein secretion system[J]. The Journal of Cell Biology, 2013, 201(4): 631-640. |
| 40 | REED E H, SCHUSTER B S, GOOD M C, et al. SPLIT: stable protein coacervation using a light induced transition[J]. ACS Synthetic Biology, 2020, 9(3): 500-507. |
| 41 | ZHANG W, LOHMAN A W, ZHURAVLOVA Y, et al. Optogenetic control with a photocleavable protein, PhoCl[J]. Nature Methods, 2017, 14(4): 391-394. |
| 42 | ZHANG W M, ZHAO G H, LUO Z Q, et al. Engineering the ribosomal DNA in a megabase synthetic chromosome[J]. Science, 2017, 355(6329): eaaf3981. |
| 43 | ABDO H, CALVO-ENRIQUE L, LOPEZ J M, et al. Specialized cutaneous Schwann cells initiate pain sensation[J]. Science, 2019, 365(6454): 695-699. |
| 44 | ALEXANDRE M T A, ARENTS J C, VAN GRONDELLE R, et al. A base-catalyzed mechanism for dark state recovery in the Avena sativa phototropin-1 LOV2 domain[J]. Biochemistry, 2007, 46(11): 3129-3137. |
| 45 | HEINTZEN C, LOROS J, DUNLAP J C. The PAS protein VIVID defines a clock-associated feedback loop that represses light input, modulates gating, and regulates clock resetting[J]. Cell, 2001, 104: 453-464. |
| 46 | FREDDOLINO P L, DITTRICH M, SCHULTEN K. Dynamic switching mechanisms in LOV1 and LOV2 domains of plant phototropins[J]. Biophysical Journal, 2006, 91(10): 3630-3639. |
| 47 | HARPER S M, NEIL L C, GARDNER K H. Structural basis of a phototropin light switch[J]. Science, 2003, 301(5639): 1541-1544. |
| 48 | HE L, TAN P, ZHU L, et al. Circularly permuted LOV2 as a modular photoswitch for optogenetic engineering[J]. Nature Chemical Biology, 2021, 17(8): 915-923. |
| 49 | KAWANO F, SUZUKI H, FURUYA A, et al. Engineered pairs of distinct photoswitches for optogenetic control of cellular proteins[J]. Nature Communications, 2015, 6: 6256. |
| 50 | NAGEL G, OLLIG D, FUHRMANN M, et al. Channelrhodopsin-1: a light-gated proton channel in green algae[J]. Science, 2002, 296(5577): 2395-2398. |
| 51 | NAGEL G, SZELLAS T, HUHN W, et al. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(24): 13940-13945 |
| 52 | PARK H, KIM N Y, LEE S, et al. Optogenetic protein clustering through fluorescent protein tagging and extension of CRY2[J]. Nature Communications, 2017, 8: 30. |
| 53 | TASLIMI A, VRANA J D, CHEN D, et al. An optimized optogenetic clustering tool for probing protein interaction and function[J]. Nature Communications, 2014, 5: 4925. |
| 54 | KENNEDY M J, HUGHES R M, PETEYA L A, et al. Rapid blue-light-mediated induction of protein interactions in living cells[J]. Nature Methods, 2010, 7(12): 973-975. |
| 55 | MOTTA-MENA L B, READE A, MALLORY M J, et al. An optogenetic gene expression system with rapid activation and deactivation kinetics[J]. Nature Chemical Biology, 2014, 10(3): 196-202. |
| 56 | WANG R, YANG Z G, LUO J R, et al. B12-dependent photoresponsive protein hydrogels for controlled stem cell/protein release[J]. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(23): 5912-5917. |
| 57 | JOST M, FERNÁNDEZ-ZAPATA J, POLANCO M C, et al. Structural basis for gene regulation by a B12-dependent photoreceptor[J]. Nature, 2015, 526(7574): 536-541. |
| 58 | MANSOURI M, HUSSHERR M D, STRITTMATTER T, et al. Smart-watch-programmed green-light-operated percutaneous control of therapeutic transgenes[J]. Nature Communications, 2021, 12: 3388. |
| 59 | XU D D, RICKEN J, WEGNER S V. Turning cell adhesions ON or OFF with high spatiotemporal precision using the green light responsive protein CarH[J]. Chemistry, 2020, 26(44): 9859-9863. |
| 60 | HABUCHI S, ANDO R, DEDECKER P, et al. Reversible single-molecule photoswitching in the GFP-like fluorescent protein Dronpa[J]. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(27): 9511-9516. |
| 61 | DAY R N, DAVIDSON M W. The fluorescent protein palette: tools for cellular imaging[J]. Chemical Society Reviews, 2009, 38(10): 2887-2921. |
| 62 | ZHOU X X, CHUNG H K, LAM A J, et al. Optical control of protein activity by fluorescent protein domains[J]. Science, 2012, 338(6108): 810-814. |
| 63 | ZHOU X X, FAN L Z, LI P P, et al. Optical control of cell signaling by single-chain photoswitchable kinases[J]. Science, 2017, 355(6327): 836-842. |
| 64 | TOETTCHER J E, WEINER O D, LIM W A. Using optogenetics to interrogate the dynamic control of signal transmission by the Ras/Erk module[J]. Cell, 2013, 155(6): 1422-1434. |
| 65 | OCHOA-FERNANDEZ R, SAMODELOV S L, BRANDL S M, et al. Optogenetics in plants: red/far-red light control of gene expression[J]. Methods in Molecular Biology, 2016, 1408: 125-139. |
| 66 | WU J L, WANG M Y, YANG X P, et al. A non-invasive far-red light-induced split-Cre recombinase system for controllable genome engineering in mice[J]. Nature Communications, 2020, 11: 3708. |
| 67 | YU G L, ZHANG M L, GAO L, et al. Far-red light-activated human islet-like designer cells enable sustained fine-tuned secretion of insulin for glucose control[J]. Molecular Therapy, 2022, 30(1): 341-354. |
| 68 | REDCHUK T A, OMELINA E S, CHERNOV K G, et al. Near-infrared optogenetic pair for protein regulation and spectral multiplexing[J]. Nature Chemical Biology, 2017, 13(6): 633-639. |
| 69 | REDCHUK T A, KARASEV M M, VERKHUSHA P V, et al. Optogenetic regulation of endogenous proteins[J]. Nature Communications, 2020, 11: 605. |
| 70 | SOROKINA O, KAPUS A, TERECSKEI K, et al. A switchable light-input, light-output system modelled and constructed in yeast[J]. Journal of Biological Engineering, 2009, 3: 15. |
| 71 | ZHOU Y, KONG D Q, WANG X Y, et al. A small and highly sensitive red/far-red optogenetic switch for applications in mammals[J]. Nature Biotechnology, 2022, 40(2): 262-272. |
| 72 | CHRISTIE J M, ARVAI A S, BAXTER K J, et al. Plant UVR8 photoreceptor senses UV-B by tryptophan-mediated disruption of cross-dimer salt bridges[J]. Science, 2012, 335(6075): 1492-1496. |
| 73 | SARRIS M, OLEKHNOVITCH R, BOUSSO P. Manipulating leukocyte interactions in vivo through optogenetic chemokine release[J]. Blood, 2016, 127(23): e35-e41. |
| 74 | LEE J, NATARAJAN M, NASHINE V C, et al. Surface sites for engineering allosteric control in proteins[J]. Science, 2008, 322(5900): 438-442. |
| 75 | PETER E, DICK B, BAEURLE S A. Mechanism of signal transduction of the LOV2-Jα photosensor from Avena sativa [J]. Nature Communications, 2010, 1: 122. |
| 76 | LUNGU O I, HALLETT R A, CHOI E J, et al. Designing photoswitchable peptides using the AsLOV2 domain[J]. Chemistry & Biology, 2012, 19(4): 507-517. |
| 77 | HARTWICK A T E, BRAMLEY J R, YU J N, et al. Light-evoked calcium responses of isolated melanopsin-expressing retinal ganglion cells[J]. The Journal of Neuroscience: the Official Journal of the Society for Neuroscience, 2007, 27(49): 13468-13480. |
| 78 | BAASKE J, GONSCHOREK P, ENGESSER R, et al. Dual-controlled optogenetic system for the rapid down-regulation of protein levels in mammalian cells[J]. Scientific Reports, 2018, 8: 15024. |
| 79 | VAN HAREN J, CHARAFEDDINE R A, ETTINGER A, et al. Local control of intracellular microtubule dynamics by EB1 photodissociation[J]. Nature Cell Biology, 2018, 20(3): 252-261. |
| 80 | BUBECK F, HOFFMANN M D, HARTEVELD Z, et al. Engineered anti-CRISPR proteins for optogenetic control of CRISPR-Cas9[J]. Nature Methods, 2018, 15(11): 924-927. |
| 81 | RENICKE C, SCHUSTER D, USHERENKO S, et al. A LOV2 domain-based optogenetic tool to control protein degradation and cellular function[J]. Chemistry & Biology, 2013, 20(4): 619-626. |
| 82 | GIL A A, CARRASCO-LÓPEZ C, ZHU L Y, et al. Optogenetic control of protein binding using light-switchable nanobodies[J]. Nature Communications, 2020, 11: 4044. |
| 83 | KAWANO F, OKAZAKI R, YAZAWA M, et al. A photoactivatable Cre-loxP recombination system for optogenetic genome engineering[J]. Nature Chemical Biology, 2016, 12(12): 1059-1064. |
| 84 | FOIGHT G W, WANG Z Z, WEI C T, et al. Multi-input chemical control of protein dimerization for programming graded cellular responses[J]. Nature Biotechnology, 2019, 37(10): 1209-1216. |
| 85 | VISWANATHAN R, HARTMANN J, PALLARES CARTES C, et al. Desensitisation of Notch signalling through dynamic adaptation in the nucleus[J]. The EMBO Journal, 2021, 40(18): e107245. |
| 86 | YUMEREFENDI H, WANG H, DICKINSON D J, et al. Light-dependent cytoplasmic recruitment enhances the dynamic range of a nuclear import photoswitch[J]. ChemBioChem, 2018, 19(12): 1319-1325. |
| 87 | GRIFFIN E A, STAKNIS D, WEITZ C J. Light-independent role of CRY1 and CRY2 in the mammalian circadian clock[J]. Science, 1999, 286(5440): 768-771. |
| 88 | AIRAN R D, THOMPSON K R, FENNO L E, et al. Temporally precise in vivo control of intracellular signalling[J]. Nature, 2009, 458(7241): 1025-1029. |
| 89 | ZHANG F, WANG L P, BRAUNER M, et al. Multimodal fast optical interrogation of neural circuitry[J]. Nature, 2007, 446(7136): 633-639. |
| 90 | GRADINARU V, ZHANG F, RAMAKRISHNAN C, et al. Molecular and cellular approaches for diversifying and extending optogenetics[J]. Cell, 2010, 141(1): 154-165. |
| 91 | ZHAO S L, CUNHA C, ZHANG F, et al. Improved expression of halorhodopsin for light-induced silencing of neuronal activity[J]. Brain Cell Biology, 2008, 36(1/2/3/4): 141-154. |
| 92 | HUGHES R M, BOLGER S, TAPADIA H, et al. Light-mediated control of DNA transcription in yeast[J]. Methods, 2012, 58(4): 385-391. |
| 93 | PIATKEVICH K D, SUBACH F V, VERKHUSHA V V. Engineering of bacterial phytochromes for near-infrared imaging, sensing, and light-control in mammals[J]. Chemical Society Reviews, 2013, 42(8): 3441-3452. |
| 94 | TSCHOWRI N, SCHUMACHER M A, SCHLIMPERT S, et al. Tetrameric c-di-GMP mediates effective transcription factor dimerization to control Streptomyces development[J]. Cell, 2014, 158(5): 1136-1147. |
| 95 | BUSH M J, TSCHOWRI N, SCHLIMPERT S, et al. C-di-GMP signalling and the regulation of developmental transitions in streptomycetes[J]. Nature Reviews Microbiology, 2015, 13(12): 749-760. |
| 96 | RYU M H, GOMELSKY M. Near-infrared light responsive synthetic c-di-GMP module for optogenetic applications[J]. ACS Synthetic Biology, 2014, 3(11): 802-810. |
| 97 | ASH C, DUBEC M, DONNE K, et al. Effect of wavelength and beam width on penetration in light-tissue interaction using computational methods[J]. Lasers in Medical Science, 2017, 32(8): 1909-1918. |
| 98 | CRICK F H. Thinking about the brain[J]. Scientific American, 1979, 241(3): 219-232. |
| 99 | DEISSEROTH K, FENG G P, MAJEWSKA A K, et al. Next-generation optical technologies for illuminating genetically targeted brain circuits[J]. The Journal of Neuroscience: the Official Journal of the Society for Neuroscience, 2006, 26(41): 10380-10386. |
| 100 | ADAMANTIDIS A R, ZHANG F, DE LECEA L, et al. Optogenetics: opsins and optical interfaces in neuroscience[J]. Cold Spring Harbor Protocols, 2014, 2014(8): 815-822. |
| 101 | BURN D J, TRÖSTER A I. Neuropsychiatric complications of medical and surgical therapies for Parkinson's disease[J]. Journal of Geriatric Psychiatry and Neurology, 2004, 17(3): 172-180. |
| 102 | LIU A L, VÖRÖSLAKOS M, KRONBERG G, et al. Immediate neurophysiological effects of transcranial electrical stimulation[J]. Nature Communications, 2018, 9: 5092. |
| 103 | ZEMELMAN B V, LEE G A, NG M, et al. Selective photostimulation of genetically ChARGed neurons[J]. Neuron, 2002, 33(1): 15-22. |
| 104 | BANGHART M, BORGES K, ISACOFF E, et al. Light-activated ion channels for remote control of neuronal firing[J]. Nature Neuroscience, 2004, 7(12): 1381-1386. |
| 105 | LIMA S Q, MIESENBöCK G. Remote control of behavior through genetically targeted photostimulation of neurons[J]. Cell, 2005, 121(1): 141-152. |
| 106 | SCHROLL C, RIEMENSPERGER T, BUCHER D, et al. Light-induced activation of distinct modulatory neurons triggers appetitive or aversive learning in Drosophila larvae[J]. Current Biology, 2006, 16(17): 1741-1747. |
| 107 | JUNG J C, MEHTA A D, AKSAY E, et al. In vivo mammalian brain imaging using one- and two-photon fluorescence microendoscopy[J]. Journal of Neurophysiology, 2004, 92(5): 3121-3133. |
| 108 | HELMCHEN F, FEE M S, TANK D W, et al. A miniature head-mounted two-photon microscope: high-resolution brain imaging in freely moving animals[J]. Neuron, 2001, 31(6): 903-912. |
| 109 | HAN X, BOYDEN E S. Multiple-color optical activation, silencing, and desynchronization of neural activity, with single-spike temporal resolution[J]. PLoS One, 2007, 2(3): e299. |
| 110 | MU D, DENG J, LIU K F, et al. A central neural circuit for itch sensation[J]. Science, 2017, 357(6352): 695-699. |
| 111 | GRADINARU V, MOGRI M, THOMPSON K R, et al. Optical deconstruction of parkinsonian neural circuitry[J]. Science, 2009, 324(5925): 354-359. |
| 112 | NGUGI A K, BOTTOMLEY C, KLEINSCHMIDT I, et al. Estimation of the burden of active and life-time epilepsy: a meta-analytic approach[J]. Epilepsia, 2010, 51(5): 883-890. |
| 113 | TØNNESEN J, SØRENSEN A T, DEISSEROTH K, et al. Optogenetic control of epileptiform activity[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(29): 12162-12167. |
| 114 | HE L, WANG L Q, ZENG H X, et al. Engineering of a bona fide light-operated calcium channel[J]. Nature Communications, 2021, 12: 164. |
| 115 | GONG X, MENDOZA-HALLIDAY D, TING J T, et al. An ultra-sensitive step-function opsin for minimally invasive optogenetic stimulation in mice and macaques[J]. Neuron, 2020, 107(1): 38-51.e8. |
| 116 | CHEN R, GORE F, NGUYEN Q A, et al. Deep brain optogenetics without intracranial surgery[J]. Nature Biotechnology, 2021, 39(2): 161-164. |
| 117 | KIM C K, SANCHEZ M I, HOERBELT P, et al. A molecular calcium integrator reveals a striatal cell type driving aversion[J]. Cell, 2020, 183(7): 2003-2019.e16. |
| 118 | TAN P, HONG T T, CAI X L, et al. Optical control of protein delivery and partitioning in the nucleolus[J]. Nucleic Acids Research, 2022, 50(12): e69. |
| 119 | STANCULEANU D L, DANIELA Z, LAZESCU A, et al. Development of new immunotherapy treatments in different cancer types[J]. Journal of Medicine and Life, 2016, 9(3): 240-248. |
| 120 | LIZÉE G, OVERWIJK W W, RADVANYI L, et al. Harnessing the power of the immune system to target cancer[J]. Annual Review of Medicine, 2013, 64: 71-90. |
| 121 | RESTIFO N P, DUDLEY M E, ROSENBERG S A. Adoptive immunotherapy for cancer: harnessing the T cell response[J]. Nature Reviews Immunology, 2012, 12(4): 269-281. |
| 122 | KHADER S A, DIVANGAHI M, HANEKOM W, et al. Targeting innate immunity for tuberculosis vaccination[J]. The Journal of Clinical Investigation, 2019, 129(9): 3482-3491. |
| 123 | FISHER B, PACKARD B S, READ E J, et al. Tumor localization of adoptively transferred indium-111 labeled tumor infiltrating lymphocytes in patients with metastatic melanoma[J]. Journal of Clinical Oncology: Official Journal of the American Society of Clinical Oncology, 1989, 7(2): 250-261. |
| 124 | GRIFFITH K D, READ E J, CARRASQUILLO J A, et al. In vivo distribution of adoptively transferred indium-111-labeled tumor infiltrating lymphocytes and peripheral blood lymphocytes in patients with metastatic melanoma[J]. Journal of the National Cancer Institute, 1989, 81(22): 1709-1717. |
| 125 | FRIDMAN W H, PAGÈS F, SAUTÈS-FRIDMAN C, et al. The immune contexture in human tumours: impact on clinical outcome[J]. Nature Reviews Cancer, 2012, 12(4): 298-306. |
| 126 | TAN P, HE L, HAN G, et al. Optogenetic immunomodulation: shedding light on antitumor immunity[J]. Trends in Biotechnology, 2017, 35(3): 215-226. |
| 127 | XU Y X, HYUN Y M, LIM K, et al. Optogenetic control of chemokine receptor signal and T-cell migration[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(17): 6371-6376. |
| 128 | HE L, ZHANG Y W, MA G L, et al. Near-infrared photoactivatable control of Ca2+ signaling and optogenetic immunomodulation[J]. eLife, 2015, 4: e10024. |
| 129 | ZHAO B X, WANG Y C, TAN X H, et al. An optogenetic controllable T cell system for hepatocellular carcinoma immunotherapy[J]. Theranostics, 2019, 9(7): 1837-1850. |
| 130 | ZHENG B, WANG H J, PAN H Z, et al. Near-infrared light triggered upconversion optogenetic nanosystem for cancer therapy[J]. ACS Nano, 2017, 11(12): 11898-11907. |
| 131 | KALIKI S, SHIELDS C L. Uveal melanoma: relatively rare but deadly cancer[J]. Eye, 2017, 31(2): 241-257. |
| 132 | AMARO A, GANGEMI R, PIAGGIO F, et al. The biology of uveal melanoma[J]. Cancer Metastasis Reviews, 2017, 36(1): 109-140. |
| 133 | SHAIN A H, BAGGER M M, YU R, et al. The genetic evolution of metastatic uveal melanoma[J]. Nature Genetics, 2019, 51(7): 1123-1130. |
| 134 | ZHANG M L, LIN X, ZHANG J P, et al. Blue light-triggered optogenetic system for treating uveal melanoma[J]. Oncogene, 2020, 39(10): 2118-2124. |
| 135 | HE L, HUANG Z X, HUANG K, et al. Optogenetic control of non-apoptotic cell death[J]. Advanced Science, 2021, 8(13): 2100424. |
| 136 | JACOBY E, NGUYEN S M, FOUNTAINE T J, et al. CD19 CAR immune pressure induces B-precursor acute lymphoblastic leukaemia lineage switch exposing inherent leukaemic plasticity[J]. Nature Communications, 2016, 7: 12320. |
| 137 | MAUS M V, JUNE C H. Making better chimeric antigen receptors for adoptive T-cell therapy[J]. Clinical Cancer Research, 2016, 22(8): 1875-1884. |
| 138 | FRIGAULT M J, LEE J, BASIL M C, et al. Identification of chimeric antigen receptors that mediate constitutive or inducible proliferation of T cells[J]. Cancer Immunology Research, 2015, 3(4): 356-367. |
| 139 | TOKAREW N, OGONEK J, ENDRES S, et al. Teaching an old dog new tricks: next-generation CAR T cells[J]. British Journal of Cancer, 2019, 120(1): 26-37. |
| 140 | CALIENDO F, DUKHINOVA M, SICILIANO V. Engineered cell-based therapeutics: synthetic biology meets immunology[J]. Frontiers in Bioengineering and Biotechnology, 2019, 7: 43. |
| 141 | CRUZ C R, HANLEY P J, LIU H, et al. Adverse events following infusion of T cells for adoptive immunotherapy: a 10-year experience[J]. Cytotherapy, 2010, 12(6): 743-749. |
| 142 | NGUYEN N T, HUANG K, ZENG H X, et al. Nano-optogenetic engineering of CAR T cells for precision immunotherapy with enhanced safety[J]. Nature Nanotechnology, 2021, 16(12): 1424-1434. |
| 143 | ZHOU S B, GRAVEKAMP C, BERMUDES D, et al. Tumour-targeting bacteria engineered to fight cancer[J]. Nature Reviews Cancer, 2018, 18(12): 727-743. |
| 144 | ZHANG X Y, ZHANG Y Y, ZHANG C N, et al. An injectable hydrogel co-loading with cyanobacteria and upconversion nanoparticles for enhanced photodynamic tumor therapy[J]. Colloids and Surfaces B: Biointerfaces, 2021, 201: 111640. |
| 145 | PAN H Z, LI L Y, PANG G J, et al. Engineered NIR light-responsive bacteria as anti-tumor agent for targeted and precise cancer therapy[J]. Chemical Engineering Journal, 2021, 426: 130842. |
| 146 | FLORES-MATEO G, CARRILLO-SANTISTEVE P, ELOSUA R, et al. Antioxidant enzyme activity and coronary heart disease: meta-analyses of observational studies[J]. American Journal of Epidemiology, 2009, 170(2): 135-147 |
| 147 | MOZAFFARIAN D, BENJAMIN E J, GO A S, et al. Heart disease and stroke statistics—2015 update: a report from the American Heart Association[J]. Circulation, 2015, 131(4): e29-e322. |
| 148 | PRIORI S G, BLOMSTRÖM-LUNDQVIST C, MAZZANTI A, et al. 2015 ESC Guidelines for the management of patients with ventricular arrhythmias and the prevention of sudden cardiac death: the Task Force for the Management of Patients with Ventricular Arrhythmias and the Prevention of Sudden Cardiac Death of the European Society of Cardiology (ESC)Endorsed by: Association for European Paediatric and Congenital Cardiology (AEPC)[J]. European Heart Journal, 2015, 36(41): 2793-2867. |
| 149 | COHEN J E, PURCELL B P, MACARTHUR J W, et al. A bioengineered hydrogel system enables targeted and sustained intramyocardial delivery of neuregulin, activating the cardiomyocyte cell cycle and enhancing ventricular function in a murine model of ischemic cardiomyopathy[J]. Circulation Heart Failure, 2014, 7(4): 619-626. |
| 150 | ASCHEIM D D, GELIJNS A C, GOLDSTEIN D, et al. Mesenchymal precursor cells as adjunctive therapy in recipients of contemporary left ventricular assist devices[J]. Circulation, 2014, 129(22): 2287-2296. |
| 151 | YANKEY G K, LI T L, KILIC A, et al. Regional remodeling strain and its association with myocardial apoptosis after myocardial infarction in an ovine model[J]. The Journal of Thoracic and Cardiovascular Surgery, 2008, 135(5): 991-998.e2. |
| 152 | VELAZQUEZ E J, LEE K L, DEJA M A, et al. Coronary-artery bypass surgery in patients with left ventricular dysfunction[J]. The New England Journal of Medicine, 2011, 364(17): 1607-1616. |
| 153 | COHEN J E, GOLDSTONE A B, PAULSEN M J, et al. An innovative biologic system for photon-powered myocardium in the ischemic heart[J]. Science Advances, 2017, 3(6): e1603078. |
| 154 | AMBROSI C M, BOYLE P M, CHEN K, et al. Optogenetics-enabled assessment of viral gene and cell therapy for restoration of cardiac excitability[J]. Scientific Reports, 2015, 5: 17350. |
| 155 | KOLOSSOV E, BOSTANI T, ROELL W, et al. Engraftment of engineered ES cell-derived cardiomyocytes but not BM cells restores contractile function to the infarcted myocardium[J]. The Journal of Experimental Medicine, 2006, 203(10): 2315-2327. |
| 156 | BRUEGMANN T, MALAN D, HESSE M, et al. Optogenetic control of heart muscle in vitro and in vivo [J]. Nature Methods, 2010, 7(11): 897-900. |
| 157 | NUSSINOVITCH U, GEPSTEIN L. Optogenetics for in vivo cardiac pacing and resynchronization therapies[J]. Nature Biotechnology, 2015, 33(7): 750-754. |
| 158 | YU L L, , ZHOU L P, CAO G, et al. Optogenetic modulation of cardiac sympathetic nerve activity to prevent ventricular arrhythmias[J]. Journal of the American College of Cardiology, 2017, 70(22): 2778-2790. |
| 159 | TILLEY D G. G protein-dependent and G protein-independent signaling pathways and their impact on cardiac function[J]. Circulation Research, 2011, 109(2): 217-230. |
| 160 | KOCKSKÄMPER J, ZIMA A V, RODERICK H L, et al. Emerging roles of inositol 1, 4, 5-trisphosphate signaling in cardiac myocytes[J]. Journal of Molecular and Cellular Cardiology, 2008, 45(2): 128-147. |
| 161 | BEIERT T, BRUEGMANN T, SASSE P. Optogenetic activation of Gq signalling modulates pacemaker activity of cardiomyocytes[J]. Cardiovascular Research, 2014, 102(3): 507-516. |
| 162 | BOJAR D, SCHELLER L, HAMRI G C E, et al. Caffeine-inducible gene switches controlling experimental diabetes[J]. Nature Communications, 2018, 9: 2318. |
| 163 | XIE M Q, YE H F, WANG H, et al. β-Cell-mimetic designer cells provide closed-loop glycemic control[J]. Science, 2016, 354(6317): 1296-1301. |
| 164 | LI T, CHEN X J, QIAN Y J, et al. A synthetic BRET-based optogenetic device for pulsatile transgene expression enabling glucose homeostasis in mice[J]. Nature Communications, 2021, 12: 615. |
| 165 | REINBOTHE T M, SAFI F, AXELSSON A S, et al. Optogenetic control of insulin secretion in intact pancreatic islets with β-cell-specific expression of Channelrhodopsin-2[J]. Islets, 2014, 6(1): e28095. |
| 166 | KUSHIBIKI T, OKAWA S, HIRASAWA T, et al. Optogenetic control of insulin secretion by pancreatic β-cells in vitro and in vivo [J]. Gene Therapy, 2015, 22(7): 553-559. |
| 167 | BELKAID Y, HAND T W. Role of the microbiota in immunity and inflammation[J]. Cell, 2014, 157(1): 121-141. |
| 168 | DURACK J, LYNCH S V. The gut microbiome: relationships with disease and opportunities for therapy[J]. The Journal of Experimental Medicine, 2019, 216(1): 20-40. |
| 169 | ZHU W H, WINTER M G, BYNDLOSS M X, et al. Precision editing of the gut microbiota ameliorates colitis[J]. Nature, 2018, 553(7687): 208-211. |
| 170 | CUI M H, SUN T, LI S, et al. NIR light-responsive bacteria with live bio-glue coatings for precise colonization in the gut[J]. Cell Reports, 2021, 36(11): 109690. |
| 171 | CUI M H, PANG G J, ZHANG T, et al. Optotheranostic nanosystem with phone visual diagnosis and optogenetic microbial therapy for ulcerative colitis at-home care[J]. ACS Nano, 2021, 15(4): 7040-7052. |
| 172 | PAN H Z, SUN T, CUI M H, et al. Light-sensitive lactococcus lactis for microbe-gut-brain axis regulating via upconversion optogenetic micro-nano system[J]. ACS Nano, 2022, 16(4): 6049-6063. |
| 173 | COMMITTEE ON SCIENCE T, LAW, POLICY, et al. The National Academies Collection: Reports funded by National Institutes of Health [M]. //OLSON S. International Summit on Human Gene Editing: A Global Discussion. Washington (DC): National Academies Press, 2016. |
| 174 | NAGY A. Cre recombinase: the universal reagent for genome tailoring[J]. Genesis, 2000, 26(2): 99-109. |
| 175 | DOW L E, FISHER J, O'ROURKE K P, et al. Inducible in vivo genome editing with CRISPR-Cas9[J]. Nature Biotechnology, 2015, 33(4): 390-394. |
| 176 | ZETSCHE B, VOLZ S E, ZHANG F. A split-Cas9 architecture for inducible genome editing and transcription modulation[J]. Nature Biotechnology, 2015, 33(2): 139-142. |
| 177 | DAVIS K M, PATTANAYAK V, THOMPSON D B, et al. Small molecule-triggered Cas9 protein with improved genome-editing specificity[J]. Nature Chemical Biology, 2015, 11(5): 316-318. |
| 178 | NGUYEN D P, MIYAOKA Y, GILBERT L A, et al. Ligand-binding domains of nuclear receptors facilitate tight control of split CRISPR activity[J]. Nature Communications, 2016, 7: 12009. |
| 179 | PAN Y C, YANG J J, LUAN X W, et al. Near-infrared upconversion-activated CRISPR-Cas9 system: a remote-controlled gene editing platform[J]. Science Advances, 2019, 5(4): eaav7199. |
| 180 | GILBERT L A, LARSON M H, MORSUT L, et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes[J]. Cell, 2013, 154(2): 442-451. |
| 181 | QI L S, LARSON M H, GILBERT L A, et al. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression[J]. Cell, 2013, 152(5): 1173-1183. |
| 182 | NIHONGAKI Y, YAMAMOTO S, KAWANO F, et al. CRISPR-cas9-based photoactivatable transcription system[J]. Chemistry & Biology, 2015, 22(2): 169-174. |
| 183 | ZETSCHE B, GOOTENBERG J S, ABUDAYYEH O O, et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system[J]. Cell, 2015, 163(3): 759-771. |
| 184 | KLEINSTIVER B P, SOUSA A A, WALTON R T, et al. Engineered CRISPR-Cas12a variants with increased activities and improved targeting ranges for gene, epigenetic and base editing[J]. Nature Biotechnology, 2019, 37(3): 276-282. |
| 185 | KIM D, KIM J, HUR J K, et al. Genome-wide analysis reveals specificities of Cpf1 endonucleases in human cells[J]. Nature Biotechnology, 2016, 34(8): 863-868. |
| 186 | KLEINSTIVER B P, TSAI S Q, PREW M S, et al. Genome-wide specificities of CRISPR-Cas Cpf1 nucleases in human cells[J]. Nature Biotechnology, 2016, 34(8): 869-874. |
| 187 | NIHONGAKI Y, OTABE T, UEDA Y, et al. A split CRISPR-Cpf1 platform for inducible genome editing and gene activation[J]. Nature Chemical Biology, 2019, 15(9): 882-888. |
| 188 | WANG X Y, DONG K L, KONG D Q, et al. A far-red light-inducible CRISPR-Cas12a platform for remote-controlled genome editing and gene activation[J]. Science Advances, 2021, 7(50): eabh2358. |
| 189 | WEBER A M, KAISER J, ZIEGLER T, et al. A blue light receptor that mediates RNA binding and translational regulation[J]. Nature Chemical Biology, 2019, 15(11): 1085-1092. |
| 190 | PILSL S, MORGAN C, CHOUKEIFE M, et al. Optoribogenetic control of regulatory RNA molecules[J]. Nature Communications, 2020, 11: 4825. |
| 191 | LIU R M, YANG J, YAO J, et al. Optogenetic control of RNA function and metabolism using engineered light-switchable RNA-binding proteins[J]. Nature Biotechnology, 2022, 40(5): 779-786. |
| 192 | VALE R D. The molecular motor toolbox for intracellular transport[J]. Cell, 2003, 112(4): 467-480. |
| 193 | WILLIAMSON R E. Organelle movements along actin filaments and microtubules[J]. Plant Physiology, 1986, 82(3): 631-634. |
| 194 | PERICO C, SPARKES I. Plant organelle dynamics: cytoskeletal control and membrane contact sites[J]. New Phytologist, 2018, 220(2): 381-394. |
| 195 | BAAS P W, DEITCH J S, BLACK M M, et al. Polarity orientation of microtubules in hippocampal neurons: uniformity in the axon and nonuniformity in the dendrite[J]. Proceedings of the National Academy of Sciences of the United States of America, 1988, 85(21): 8335-8339. |
| 196 | CONDE C, CÁCERES A. Microtubule assembly, organization and dynamics in axons and dendrites[J]. Nature Reviews Neuroscience, 2009, 10(5): 319-332. |
| 197 | DUAN L T, CHE D, ZHANG K, et al. Optogenetic control of molecular motors and organelle distributions in cells[J]. Chemistry & Biology, 2015, 22(5): 671-682. |
| 198 | VAN BERGEIJK P, ADRIAN M, HOOGENRAAD C C, et al. Optogenetic control of organelle transport and positioning[J]. Nature, 2015, 518(7537): 111-114. |
| 199 | FISCHBACH M A, BLUESTONE J A, LIM W A. Cell-based therapeutics: the next pillar of medicine[J]. Science Translational Medicine, 2013, 5(179): 179ps7. |
| 200 | FOLCHER M, OESTERLE S, ZWICKY K, et al. Mind-controlled transgene expression by a wireless-powered optogenetic designer cell implant[J]. Nature Communications, 2014, 5: 5392. |
| 201 | JENAL U, MALONE J. Mechanisms of cyclic-di-GMP signaling in bacteria[J]. Annual Review of Genetics, 2006, 40: 385-407. |
| 202 | BURDETTE D L, MONROE K M, SOTELO-TROHA K, et al. STING is a direct innate immune sensor of cyclic di-GMP[J]. Nature, 2011, 478(7370): 515-518. |
| 203 | GOMELSKY M. Photoactivated cells link diagnosis and therapy[J]. Science Translational Medicine, 2017, 9(387): eaan3936. |
| 204 | MICKLE A D, WON S M, NOH K N, et al. A wireless closed-loop system for optogenetic peripheral neuromodulation[J]. Nature, 2019, 565(7739): 361-365. |
| 205 | KATHE C, MICHOUD F, SCHÖNLE P, et al. Wireless closed-loop optogenetics across the entire dorsoventral spinal cord in mice[J]. Nature Biotechnology, 2022, 40(2): 198-208. |
| 206 | BATABYAL T, BRODOVSKAYA A, WILLIAMSON J, et al. A deep learning-based automated closed-loop optogenetic system for neuromodulation during seizures[EB/OL]. bioRxiv (2022-04-21)[2022-05-01]. . |
| 207 | TANG J, DU Y P, LEE C A, et al. Low-intensity far-red light inhibits early lesions that contribute to diabetic retinopathy: in vivo and in vitro [J]. Investigative Ophthalmology & Visual Science, 2013, 54(5): 3681-3690. |
| 208 | ZHANG F, TZANAKAKIS E S. Amelioration of diabetes in a murine model upon transplantation of pancreatic β-cells with optogenetic control of cyclic adenosine monophosphate[J]. ACS Synthetic Biology, 2019, 8(10): 2248-2255. |
| 209 | XIE M Q, YE H F, HAMRI G C E, et al. Antagonistic control of a dual-input mammalian gene switch by food additives[J]. Nucleic Acids Research, 2014, 42(14): e116. |
| 210 | GITZINGER M, KEMMER C, FLURI D A, et al. The food additive vanillic acid controls transgene expression in mammalian cells and mice[J]. Nucleic Acids Research, 2011, 40(5): e37. |
| 211 | CHEN R, ROMERO G, CHRISTIANSEN M G, et al. Wireless magnetothermal deep brain stimulation[J]. Science, 2015, 347(6229): 1477-1480. |
| 212 | HERNÁNDEZ-MORALES M, SHANG T, CHEN J J, et al. Lipid oxidation induced by RF waves and mediated by ferritin iron causes activation of ferritin-tagged ion channels[J]. Cell Reports, 2020, 30(10): 3250-3260.e7. |
| 213 | DECKERS R, QUESSON B, ARSAUT J, et al. Image-guided, noninvasive, spatiotemporal control of gene expression[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(4): 1175-1180. |
| 214 | PAN Y J, YOON S, SUN J, et al. Mechanogenetics for the remote and non-invasive control of cancer immunotherapy[J]. The FASEB Journal, 2018, 32: 674.24. |
| 215 | SHKARINA K, HASEL DE CARVALHO E, SANTOS J C, et al. Optogenetic activators of apoptosis, necroptosis, and pyroptosis[J]. The Journal of Cell Biology, 2022, 221(6): e202109038. |
| 216 | YIZHAR O, FENNO L E, DAVIDSON T J, et al. Optogenetics in neural systems[J]. Neuron, 2011, 71(1): 9-34 |
| 217 | OWEN S F, LIU M H, KREITZER A C. Thermal constraints on in vivo optogenetic manipulations[J]. Nature Neuroscience, 2019, 22(7): 1061-1065. |
| [1] | 高歌, 边旗, 王宝俊. 合成基因线路的工程化设计研究进展与展望[J]. 合成生物学, 2025, 6(1): 45-64. |
| [2] | 李冀渊, 吴国盛. 合成生物学视域下有机体的两种隐喻[J]. 合成生物学, 2025, 6(1): 190-202. |
| [3] | 焦洪涛, 齐蒙, 邵滨, 蒋劲松. DNA数据存储技术的法律治理议题[J]. 合成生物学, 2025, 6(1): 177-189. |
| [4] | 唐兴华, 陆钱能, 胡翌霖. 人类世中对合成生物学的哲学反思[J]. 合成生物学, 2025, 6(1): 203-212. |
| [5] | 郑益坤, 郑婕, 胡国鹏. 光遗传学工具在学习记忆中的应用研究[J]. 合成生物学, 2025, 6(1): 87-104. |
| [6] | 徐怀胜, 石晓龙, 刘晓光, 徐苗苗. DNA存储的关键技术:编码、纠错、随机访问与安全性[J]. 合成生物学, 2025, 6(1): 157-176. |
| [7] | 石婷, 宋展, 宋世怡, 张以恒. 体外生物转化(ivBT):生物制造的新前沿[J]. 合成生物学, 2024, 5(6): 1437-1460. |
| [8] | 柴猛, 王风清, 魏东芝. 综合利用木质纤维素生物转化合成有机酸[J]. 合成生物学, 2024, 5(6): 1242-1263. |
| [9] | 邵明威, 孙思勉, 杨时茂, 陈国强. 基于极端微生物的生物制造[J]. 合成生物学, 2024, 5(6): 1419-1436. |
| [10] | 陈雨, 张康, 邱以婧, 程彩云, 殷晶晶, 宋天顺, 谢婧婧. 微生物电合成技术转化二氧化碳研究进展[J]. 合成生物学, 2024, 5(5): 1142-1168. |
| [11] | 郑皓天, 李朝风, 刘良叙, 王嘉伟, 李恒润, 倪俊. 负碳人工光合群落的设计、优化与应用[J]. 合成生物学, 2024, 5(5): 1189-1210. |
| [12] | 夏孔晨, 徐维华, 吴起. 光酶催化混乱性反应的研究进展[J]. 合成生物学, 2024, 5(5): 997-1020. |
| [13] | 陈子苓, 向阳飞. 类器官技术与合成生物学协同研究进展[J]. 合成生物学, 2024, 5(4): 795-812. |
| [14] | 蔡冰玉, 谭象天, 李伟. 合成生物学在干细胞工程化改造中的研究进展[J]. 合成生物学, 2024, 5(4): 782-794. |
| [15] | 谢皇, 郑义蕾, 苏依婷, 阮静怡, 李永泉. 放线菌聚酮类化合物生物合成体系重构研究进展[J]. 合成生物学, 2024, 5(3): 612-630. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
