|
||
|
Halogenases in Biocatalysis: Advances in Mechanism Elucidation, Directed Evolution, and Green Manufacturing
Synthetic Biology Journal
DOI: 10.12211/2096-8280.2024-091
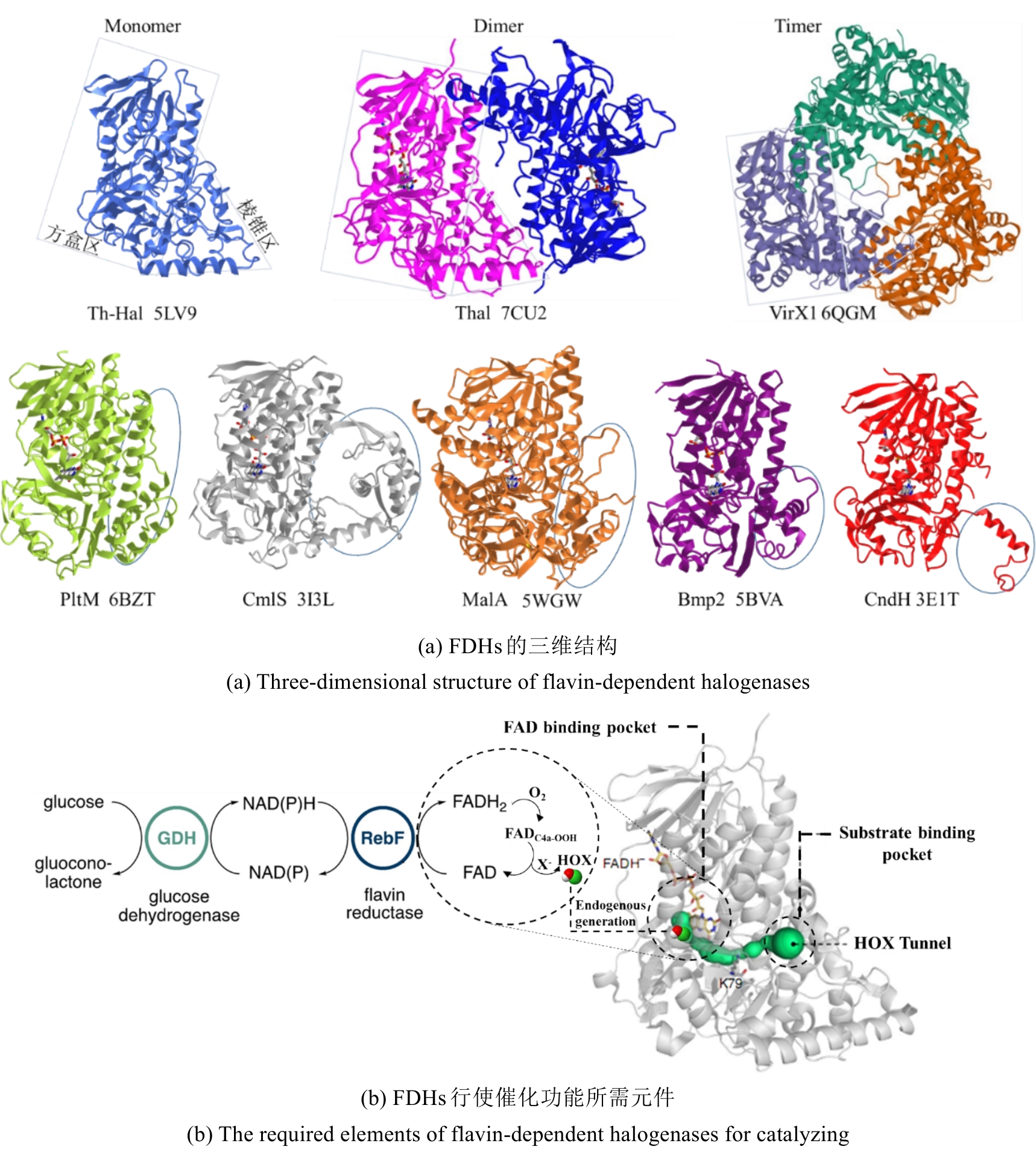
Fig. 6
The typical structure and catalytic elements of flavin-dependent halogenases
Extracts from the Article
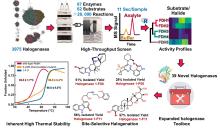
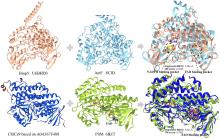
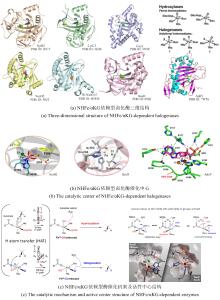
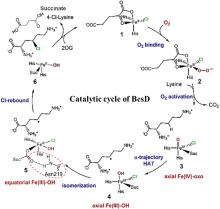
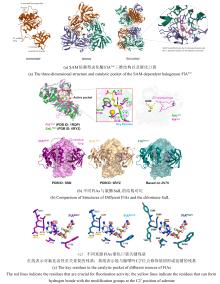
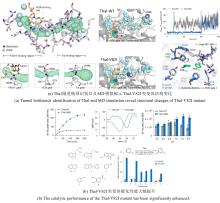
从荧光假单胞菌(Pseudomonas fluorescens)中分离的色氨酸7-卤化酶PrnA是FDHs家族的第一个成员,其X射线单晶结构也是首个被解析和发表的[61]。此后,更多FDHs被鉴定并解析,关键结构特征逐渐显现[4,17,22-23]。现有的FDHs可以根据蛋白亚基的数量分为单体(CmlS、Mpy16、PltM)、二聚体(PrnA、RebH、Thal)和三聚体(VirX1)卤化酶。在结构上,FDHs大多由两个亚结构域组成,形态上呈现为方盒区和棱锥区[图6(a)]。其中,结合黄素辅因子的方盒区高度保守,而与底物结合相关的棱锥区则变化多样[图6(a)]。FDHs的结构多样性使其适用于多种底物,比如PltM偏好卤化酚类和苯胺化合物[62],Bmp2特异性地将溴转移到吡咯类底物上[63],而CmlS主要催化脂肪族底物[64]。这些例子证明了自然界中结构决定功能的普遍规律。
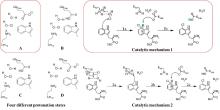
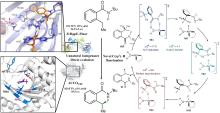
所有已知的FDHs都需要FADH2作为辅助因子[图6(b)]。FDHs的辅因子结合口袋和催化口袋分别开口于方盒区的两侧,而两个功能口袋一般通过包埋在酶内部的隧道相连,隧道长度约为10?。FADH2结合在酶的特异位点(GxGxxG)上,并与氧气反应生成黄素过氧化氢中间体(FADC4a-OOH);随后,FADC4a-OOH与卤素离子X-反应,形成次卤酸(HOX);HOX经过10?的连通隧道转移至酶催化口袋,由高度保守的活性位点赖氨酸(如PrnA-K79,RebH-K79,BrvH-K83,RadH-K74)的氨基进一步极化O-X键,并通过氢键作用将其引导至底物,催化卤化反应进行[4,17,22-23]。
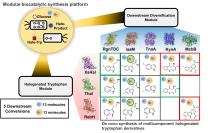
由FDHs催化机制可知,HOX的生成及转运效率是影响酶催化性能的关键因素。目前,HOX的生成严重依赖辅因子循环再生,涉及两种外源酶,三种反应以及四种辅酶因子的相互转化,反应过程复杂[图6(b)]。为提高HOX生成效率,Liu等[93]构建了一个辅酶自给自足的三酶融合体,并成功应用于L-色氨酸的连续卤化。首先,鉴定并选择来自Streptomyces pratensis的高选择性卤化酶SpFDH。然后,采用基因融合技术,将SpFDH与葡萄糖脱氢酶(GDH)和黄素还原酶(FR)融合,形成了一个三酶融合体,实现辅酶自给自足,6-氯色氨酸产率提高了1.46倍。随后,作者团队还开发了一种将三酶融合体固定化并连接到连续催化装置上的连续卤化生物工艺,反应12小时后6-氯色氨酸转化率可达97.6%。该研究为多酶级联催化制备卤代色氨酸提供了一种简明策略。
与HPLC、MS和NMR等上机检测方法相比,荧光信号能够实现快速、高通量、可视化检测从而提高筛选效率.Reed等[ Fluorescence-based screens for engineering enzymes linked to halogenated tryptophan 1 2024 ... 与HPLC、MS和NMR等上机检测方法相比,荧光信号能够实现快速、高通量、可视化检测从而提高筛选效率.Reed等[ Crystallization and X-ray diffraction of a halogenating enzyme, tryptophan 7-halogenase, from Pseudomonas fluorescens 2 2004 ... 从荧光假单胞菌(Pseudomonas fluorescens)中分离的色氨酸7-卤化酶PrnA是FDHs家族的第一个成员,其X射线单晶结构也是首个被解析和发表的[
Other Images/Table from this Article
|


{kind=link}